Kamagra gibt es auch als Kautabletten, die sich schneller auflösen als normale Pillen. Manche Patienten empfinden das als angenehmer. Wer sich informieren will, findet Hinweise unter kamagra kautabletten.
Vetrade.ir
Chapter 10
Artificial Insemination in Poultry
M.R. Bakst and J.S. Dymond
Additional information is available at the end of the chapter
Artificial insemination (AI) is the manual transfer of semen into the female's vagina. Basicallyit is a two step procedure: first, collecting semen from the male [1]; and second, inseminatingthe semen into the female [2]. In poultry, depending on the objectives and goals of the farm orlaboratory, there may be intervening steps such as semen dilution, storage, and evaluation.
Artificial insemination is practiced extensively with commercial turkeys. This is primarily theresult of selective breeding for a heavier and broader-breasted commercial turkey and theconsequent inability of toms to consistently transfer semen to the hen at copulation. The broilerindustry has not adapted AI to the extent of the turkey industry but it is occasionally used inpedigree lines and in regions where labor is relatively cheap.
To grasp the magnitude of AI in the turkey industry compared to that of livestock, a hypo‐thetical flock of 500 breeder hens inseminated with 100 μL of diluted semen (1:1) twice theweek before the onset of egg production and once weekly thereafter for the 24 wk of eggproduction would entail 13,000 inseminations using 650 mL of semen. It should be apparentwith these numbers, semen collection and hen inseminations are labor intensive as each maleand female must be handled each week.
Looking back over the use of AI in the turkey industry one can safely say that in the 1960s,weekly inseminations were based on semen volume per dose using undiluted semen. In the1970s and early 1980s, breeder farms began to dilute semen and inseminate a known numberof sperm per dose. In the mid-1980s through the 1990s, hens were initially inseminated a weekbefore the onset of lay and inseminations were performed with a known number of ‘viable'sperm. Currently, while inseminating before the onset of egg production remains widelypracticed, most companies, but not all [3], have gone back to inseminating a known volumeof semen or number of sperm per dose, in the 1970s and 1980s.
2013 Bakst and Dymond; licensee InTech. This is an open access article distributed under the terms of theCreative Commons Attribution License (http://creativecommons.org/licenses/by/3.0), which permitsunrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Success in Artificial Insemination - Quality of Semen and Diagnostics Employed
In the following chapter we will review the basics of AI and fertility evaluation in poultry. To
better appreciate the biological basis of these techniques, an overview of the reproductive
biology of poultry is provided. Detailed descriptions of techniques for the collection, evalua‐
tion, dilution, and storage of poultry semen are available in a recent publication by Bakst and
Long [4]. Earlier comprehensive reviews include Lake and Stewart [5], Bakst and Wishart [6],
and Bakst and Cecil [7].
2. Reproductive biology of poultry
This section will introduce to some and review for others the strategy of avian reproduction
with emphasis on the hen. For more comprehensive reviews on reproduction in the avian male
and female see Jamieson [8].
2.1. Overview
The goal of AI is to produce a succession of fertilized eggs between successive inseminations.
To accomplish this, weekly inseminations must replenish the sperm population in the
uterovaginal junction (UVJ) sperm storage tubules (SSTs). Birds do not have an estrous cycle
that synchronizes copulation with ovulation. Alternatively, about 7-10 days before their first
ovulation, hens mate, sperm ascend the vagina and then enter the SSTs. At the onset of egg
production, individual sperm are slowly released from the SSTs, transported to the anterior
end of the oviduct, and interact with the surface of the ovum (see [9-10] for recent reviews).
Whether fertilized or not, over the next 24-26 hr the ovum is transported though the oviduct,
accruing the outer perivitelline layer (PL) in the infundibulum, the albumen in the magnum,
the shell membrane in the isthmus, and the hard shell in the uterus (also referred to as the shell
gland) before oviposition. If fertilized, the blastoderm in the first laid egg consists of
40,000-60,000 cells in the turkey and 80,000-100,000 cells in the chicken.
Ovary: In the hen only the left ovary and oviduct become functional organs. About 2-3 wk
before the onset of lay, small (less than 1 mm in diameter) white-yolk follicles begin to
accumulate yellow yolk with some being recruited into a hierarchy of maturing yellow-yolk
follicles (Figure 1). At the time of ovulation, the largest follicle, designated as F1, is ovulated.
About 17 days were necessary for the 1 mm diameter white yolk follicle to mature to a pre-
ovulatory 40 mm diameter yellow yolk follicle [11]. After the F1 follicle is ovulated, the next
largest follicle, formerly designated F2, becomes the F1 follicle and will ovulate at the beginning
of the next daily "ovulatory cycle" in 24-26 hr.
The follicular sheath surrounding the maturing oocyte consists of histologically distinct con‐
centric layers of cells: the outer serosa (germinal epithelium); the theca externa, which forms the
greatest portion of the follicle wall, provides structural support to the follicle and has steriodo‐
genic cells; the theca interna, a highly vascularized layer, which like the theca externa has ste‐
roid-producing cells (both thecal layers synthesize androgens and estrogens); and, the
granulosa cell layer, enveloping the oocyte, which is responsible for progesterone secretion and
the synthesis of the inner PL. The inner PL is homologous to the mammalian zona pellucida and
Artificial Insemination in Poultry
is a fibrous reticulum about 2 μm thick. At ovulation, only the inner PL envelops the ovum.
While there is no corpus luteum formation in birds, the thecal layers and the granulosa of the
post-ovulatory follicle (POF) produce prostaglandins [12] and progesterone, respectively
[13-16] then regress over the next 72 hr. The POF has a pocket like appearance after ovulation
(Figure 1). On the surface of the inner PL overlying the germinal disc (GD), which is a 3.5 mm di‐
ameter disc of white yolk containing the haploid pronucleus and associated organelles, are
sperm receptors. Sperm bind to the receptors overlying the GD, hydrolyze a path through the
inner PL, and are incorporated into the ovum. Polyspermy is normal in birds but only one sperm
in apposition to the female pronucleus undergoes nuclear decondensation and initiates synga‐
my, the reconstitution of the diploid number of chromosomes.
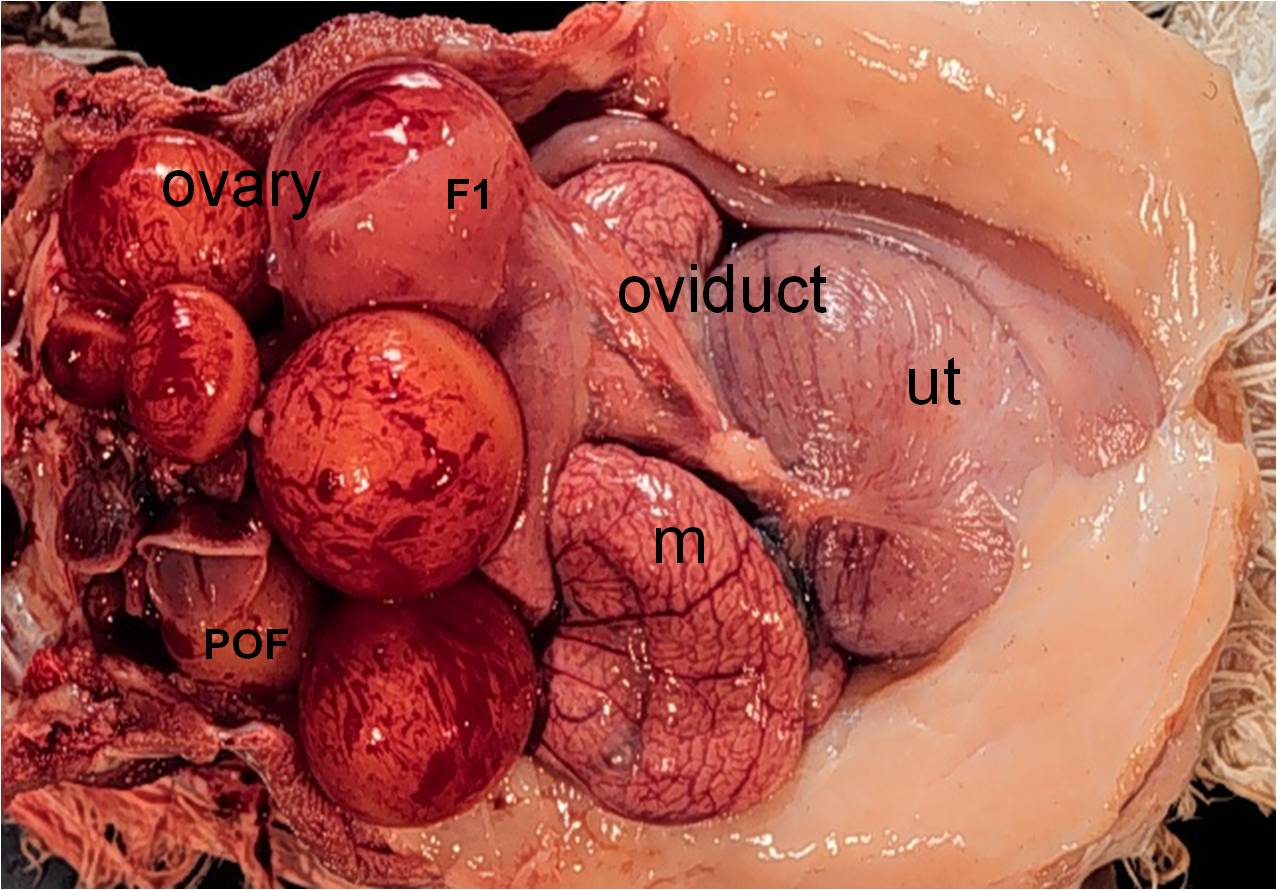
Figure 1. The ovary and oviduct of a turkey hen in egg production occupy much of the abdominal cavity. The ovarian
follicular hierarchy consisting of ovarian follicles at various stages of develop (7 maturing follicles visible in this photo‐
graph) is observed. The largest follicle, F1 follicle is the next to ovulate. The ovum ovulated about 10 hr earlier has
accrued albumen in the magnum (m), a shell membrane in the isthmus, and is observed in the uterus (ut) undergoing
shell formation. Its post-ovulatory follicular sheath (POF) appears as an open pocket. The vagina (distal to the uterus
and not visible) is embedded in connective tissue and enveloped by the abdominal fat pad.
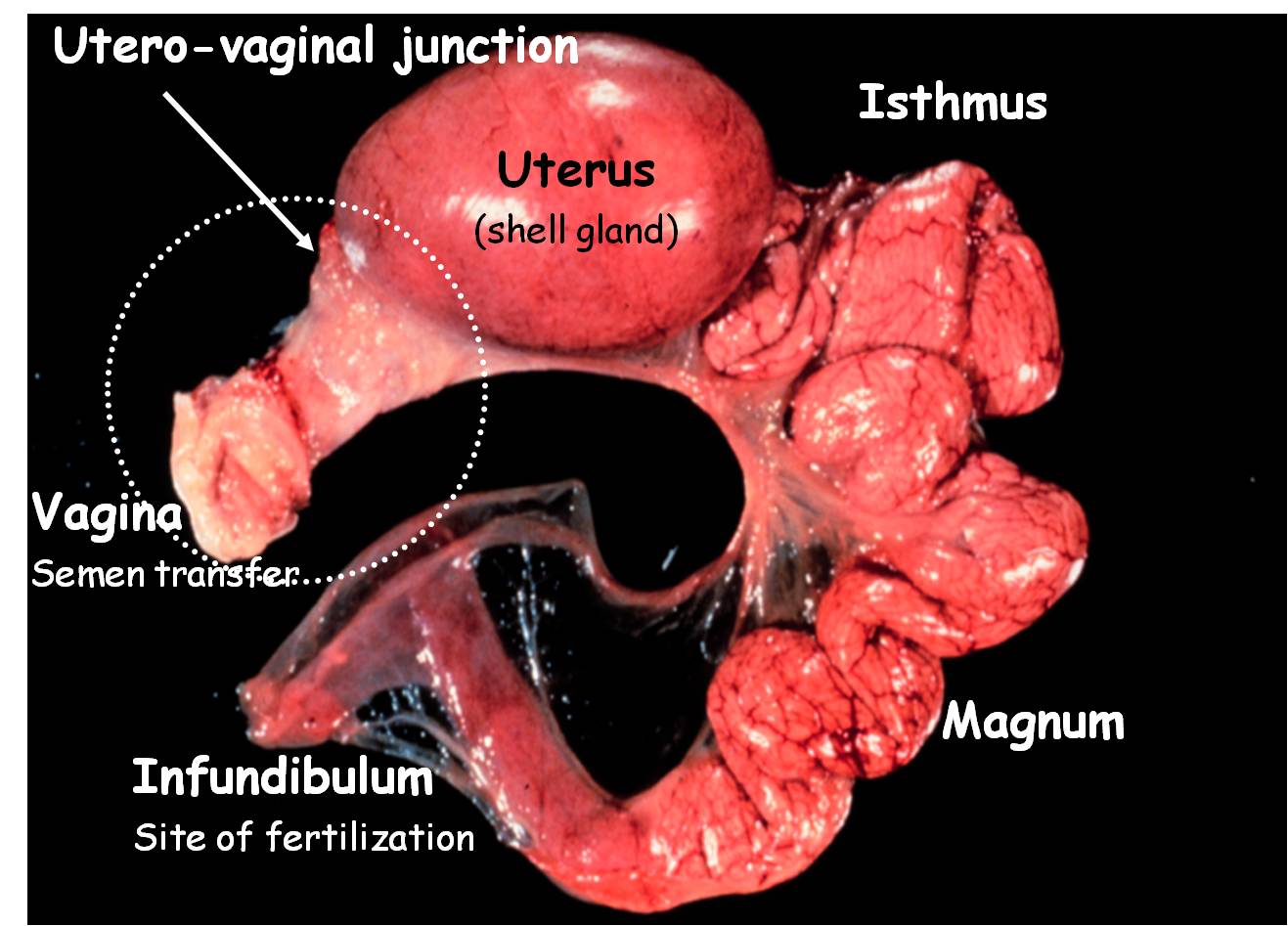
2.2. Oviduct
The mature oviduct consists of five anatomically and functionally distinct segments (Figures1 and 2): the infundibulum, which secretes an albumen-like product that forms the outer PLand prevents pathological polyspermy; the magnum, responsible for deposition of the
Success in Artificial Insemination - Quality of Semen and Diagnostics Employed
albumen proteins; the isthmus, which forms the shell membrane; the uterus (also referred toas the shell gland), a pocket-like structure that elaborates the hard-shell; and, the vagina, whichis a conduit between the uterus and cloaca for the egg-mass at oviposition and is responsiblefor sperm selection and storage following semen transfer. Interestingly, when the vagina anduterus are excised and fixed in toto and the connective tissue surrounding the vagina subse‐quently removed, the vagina appears as a coiled segment (Figure 3) [10]. This anatomy explainsthe resistance one feels when performing a vaginal insemination with a straw regardless ofthe presence or absence of an egg mass in the uterus. If inseminating a hen within 30 min afteroviposition, the connective tissue around the vagina and the smooth muscle composing thevaginal wall are flaccid. Venting (exteriorizing the vagina for placement of the inseminatingstraw) at this time may induce a partial prolapse leading to a deep insemination (closer to theUVJ) and the forfeiture of sperm selection by the vagina. Such deep inseminations areassociated with high embryo mortality, possibly due to pathological polyspermy.
The surface mucosa of each segment of the oviduct is lined with parallel, gently spiraling foldsalong the longitudinal axis. The surface epithelium lining the luminal mucosa contains varyingproportions of secretory and ciliated cells. All segments except the fimbriated region of theinfundibulum and the vagina possess sub-epithelial tubular glands that secrete componentsused in egg formation [17]. However, the anterior 2-3 cm of the vagina, an area referred to asthe UVJ (Figure 3), contains the SSTs, the primary sites of sperm storage [10] (Figure 4).
At ovulation, the ovum is grasped by the fimbriated region of the infundibulum and, if spermare present, the ovum may be fertilized within a 10-15 min interval [18]. Thereafter, infundib‐ular secretions accrue around the ovum, forming the outer PL, which acts as a barrier to furthersperm penetration. Birkhead [19] observed that the number of sperm trapped in the outer PLis positively correlated with the size of the ovum and is likewise correlated with the numberof sperm that have penetrated the inner PL. Interestingly, the sperm trapped in the outer PLretain an intact acrosome [20-21]. If fertilized, the first cleavage furrow in the GD appears 7-8hr post-ovulation, while the egg-mass is in the isthmus.
2.3. Oviductal sperm selection, transport, and storage
Following deposition in the oviduct, sperm are transported to UVJ by a combination of their
intrinsic motility and cilia beat activity [9-10, 22-23]. Within the SST lumen, sperm are either
widely spaced or oriented parallel with their heads toward the distal end of the SST (Figure
4). Sperm are apposed to, but not directly contacting the apical microvilli of the SST epithelial
cells. This spatial relationship may facilitate lipid transfer between the resident sperm and the
SST epithelial cells [24-25]. Interestingly, alkaline phosphatase, known to play a role in lipid
transfer, has been histochemically localized in the apical region of the SST epithelium [26].
The duration of sperm storage in the SSTs is species-dependent. Chickens can store sperm for
up to three weeks, whereas turkeys can maintain sperm for 10 weeks in the SST and still lay a
fertilized ovum [27-28]. This may be related to number of SSTs present in the UVJ; turkeys
have been reported to have 20,000-30,000 SSTs, while chickens have been estimated to have
Artificial Insemination in Poultry
only 5,000-13,500 [29-30]. Additionally, after several generations of selection for high fertility,chicken hens possessed increased numbers of SSTs when compared to non-selected controlhens, suggesting the number of SSTs may be positively correlated with fertility [31]. In contrast,under commercial conditions, different broiler strains exhibiting different fertility levelsrevealed similar numbers of SSTs [29].
Figure 2. The segments of the turkey oviduct with a hard-shelled egg in the uterus are observed. Sperm transferred
into the vagina undergo an intense selection process before reaching the sperm storage tubules (SSTs) localized in the
utero-vaginal junction. Sperm are slowly released from the SSTs and ascend to the infundibulum, the site of fertiliza‐
tion. In this photograph, the vagina is enveloped by connective tissue.
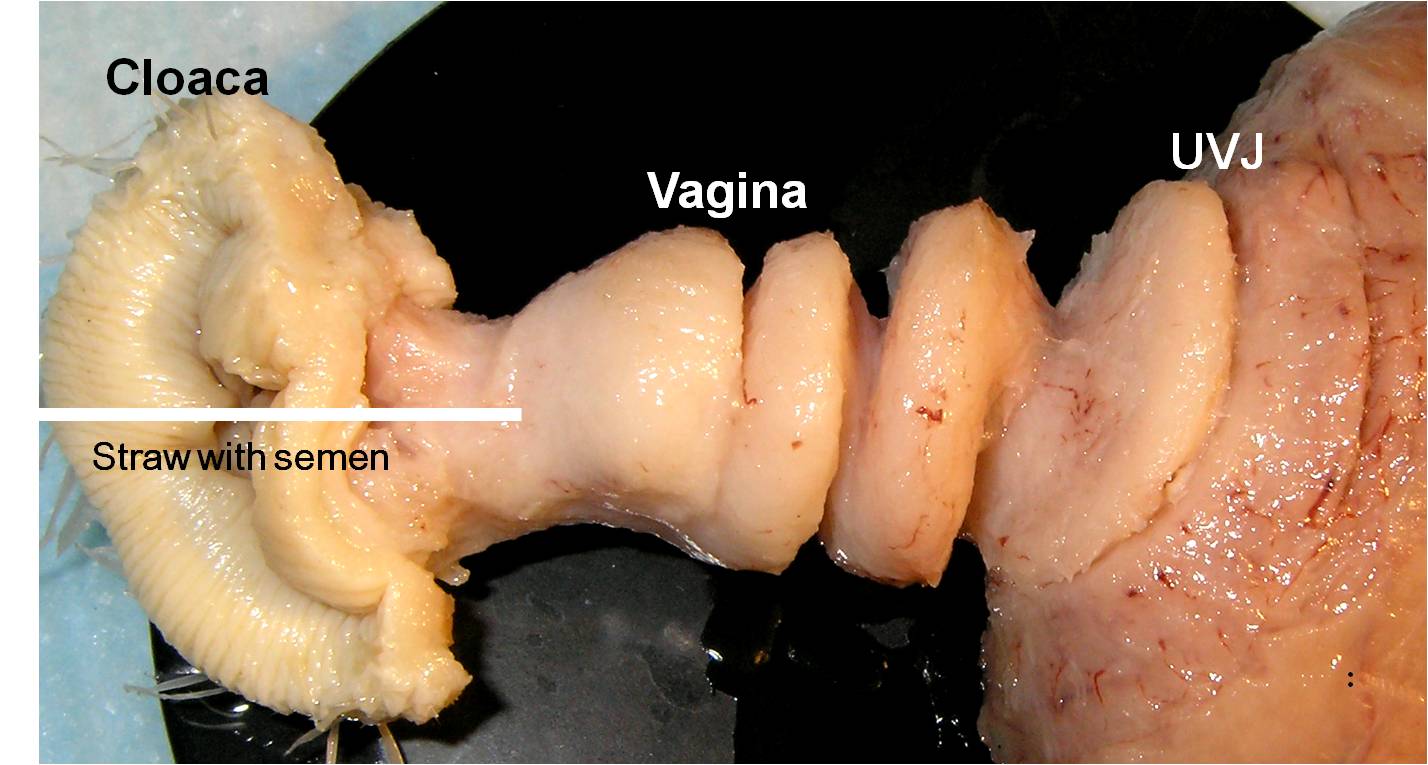
Success in Artificial Insemination - Quality of Semen and Diagnostics Employed
Figure 3. Following fixation in neutral-buffered formalin and the removal of the surrounding connective tissue, the
coiled morphology of the turkey vagina is revealed. When inseminating a hen, one should insert the straw with the
semen until resistance is felt, then release the semen as the straw is withdrawn. As observed here the resistance is due
to the coiled vaginal and not an egg mass in the uterus.
Little is known concerning the cellular and molecular mechanisms that sustain sperm withinthe SST lumen for prolonged periods of storage. These mechanisms likely involve the rever‐sible suppression of sperm motility and metabolism, protection and repair of the sperm plasmamembrane, uptake and storage of molecules to sustain sperm metabolism, and maintenanceof the SST lumen by removing by-products of sperm metabolism and degraded sperm [32-33].
It is clear the SSTs generate a discrete environment to maintain sperm viability via the influxand efflux of compounds critical for sperm survival [25, 34). While ultrastructural analysis hasrevealed only limited evidence of secretory activity [25], the identification of membrane-boundvesicles released from the apical tips of the SST epithelial cell microvilli suggests a role in themaintenance of resident sperm through lipid transfer [22, 25, 26, 32, 35, 36]. A large proportionof the sperm plasma membrane is composed of polyunsaturated fatty acids [37] that are highlysusceptible to damage induced by lipid peroxidation [37]. The peroxidation of these fatty acidsresults in increased damage to and permeability of the sperm plasma membrane [39, 40]. Acomplex system of anti-oxidation enzymes are present in the SST epithelial cells and presum‐ably interact with luminal sperm to minimize damage due to lipid peroxidation and maintainsperm membrane integrity [41]. While many metabolites required by sperm in the SSTs haveyet to be identified, increased avidin expression is apparent in SSTs relative to surroundingUVJ epithelial tissue possibly providing a means of sequestering biotin and other vitamins foruse by the SSTs or resident sperm [42-43]. Interestingly, progesterone has been shown to induceexpression of avidin in the oviduct, providing a potential link between progesterone fluctua‐tion and sperm storage in and release from the SSTs [42, 44, 45].

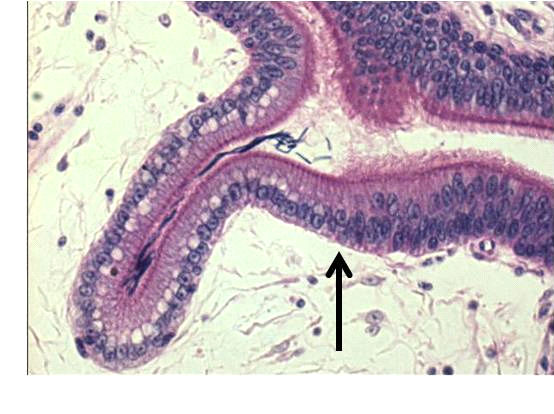
Artificial Insemination in Poultry
Figure 4. Three views of the turkey's sperm-storage tubules (SSTs) are observed. The left panel is a stereoscope image
showing the pleomorphic appearance of the SSTs. The length of the SST can be as long as 300μm. In the right panel a
hen was inseminated with sperm stained with Hoechst 33342, a nuclear fluorescent dye, the UVJ mucosa containing
SSTs was isolated, and an unfixed squash preparation was observed by dual interference contrast and fluorescence
microscopy. Sperm with fluorescing nuclei are observed in the two SST lumina. The lower-middle panel shows a histo‐
logical section of a portion of a SST containing sperm (the dense rod-like structures in the lumen are sperm nuclei. The
arrow indicates the transition between the pseudo-stratified columnar ciliated epithelium of the uterovaginal junction
and the simple columnar epithelium of the SST that is characterized in histological preparations by the supra-nuclear
vacuole.
Sperm exit the SSTs in a slow, continuous stream [46-49]; however, a stimulus cuing the egressof resident sperm from the SSTs has yet to be identified. The observations that receptors forestrogen and progesterone exist in the SSTs has led to the suggestion that these compoundsmay trigger release of resident sperm, possibly in response to hormonal cues over the courseof the ovulatory cycle[50-52]. However, an alternate theory suggests the inherent mobility ofthe sperm plays a larger role than hormonal induction in egress of sperm from SSTs [9].
Resident sperm exhibit a slow, synchronized oscillatory movement in the lumen of SSTs,suggesting the presence of a fluid current through the SST lumen [23-24]. The identification ofwater channels, known as aquaporins, in the apical epithelium of SSTs lends credence to amodel wherein motile sperm maintain their residence in the SST lumen by swimming againstthe fluid current generated via the aquaporins [53-56]. In the SST lumen, sperm retain theirmotility by fatty acid oxidation. It has been suggested the sperm membrane is the source ofthis fatty acid and that as the quality of the sperm membrane gradually decreases there is a
Success in Artificial Insemination - Quality of Semen and Diagnostics Employed
reduction of available ATP and sperm motility decreases [56]. Sperm are then swept out of the
SST lumen into the UVJ, where they encounter various stimuli enhancing their motility. These
sperm are then transported to the infundibulum, the site of fertilization [57]. Such motility-
enhancing factors may include changes in environmental pH and neuroendocrine factors such
as serotonin [58-62]. Further oxidation of sperm fatty acids, possibly sequestered from the
surround milieu, generates the energy required for sperm to respond to such motility-
enhancing factors and transcend the oviduct [9, 22, 55, 63].
Once sperm are deposited in the oviduct, several selection barriers must be overcome prior to
ascending to the infundibulum and fertilizing an ovum. This selection occurs initially in the
vagina: only highly mobile (defined as progressive movement in a viscous medium at 40oC)
sperm traverse the vagina [9]. While sperm mobility is a major factor in sperm selection in the
vagina, sperm selection is also dependent upon the glycoprotein composition of the sperm
plasma membrane. The sperm glycocalyx is highly complex and heavily sialylated and
modification of the glycocalyx results in reduced fertility and failure of the sperm to enter the
SSTs [64-67]. Interestingly, removal of membrane-associated carbohydrates did not affect
sperm entry into SSTs if sperm were inseminated directly into the UVJ or when co-incubated
with UVJ explants, suggesting the glycocalyx plays a central role in sperm transport and
selection through the vagina [64, 66, 68]. Further barriers to sperm prior participating in the
process of fertilization include sperm release from the SST and subsequent transport to the
infundibulum, and their interaction with the ovum (reviewed in [69]).
2.4. Sperm: Ovum interaction and fertilization
Given the voluminous nature of the hen's ovum and the GD relative to mammalian ova, one
must assume that yet-to-be identified factors "attract" sperm to the GD. Examination of the
electrophoretic profile of the GD and non-GD regions of the PL revealed no variation in pro‐
tein composition [70]. Furthermore, the abrogation of the preferential interaction of sperm
and the inner PL overlying the GD in vitro suggests the factors underlying the preferential
binding of sperm are not necessarily associated with the inner PL [70]. It is clear, however,
glycoproteins play a large role in the interaction between the sperm and ova, even if not di‐
rectly involved in targeting of sperm to the GD in vivo [71]. Pre-treatment of either the PL or
sperm with N-glycanases resulted in significantly decreased sperm-ovum interaction in vitro
[68, 71]. Conversely, N-linked oligosaccharides released from the inner PL by N-glycosidase
treatment could induce the acrosome reaction in sperm in vitro [72]. These findings strongly
suggest N-linked glycans, most likely terminal N-acetyl glucosamine residues, have an es‐
sential role in the sperm-ovum interaction in avian species, specifically in induction of the
acrosome reaction [68, 72].
Interaction between the sperm and inner PL results in induction of the acrosome reaction
[73]. During the acrosome reaction, the inner and outer acrosomal membranes dehisce re‐
sulting in the release of acrosin (a trypsin-like enzyme) [21, 74]. As the result of the acro‐
some reaction, sperm hydrolyze a small hole in the inner PL (Figure 5), enabling sperm to
reach the microvilli-studded surface of the ovum [21, 74]. The capacity of sperm to hydro‐
lyze and penetrate the inner PL is the biological basis for the sperm penetration assay dis‐
cussed below and next section.
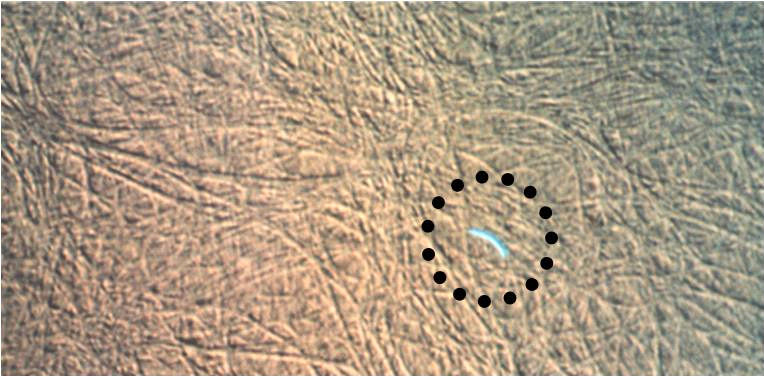
Artificial Insemination in Poultry
Figure 5. In the left panel, a turkey sperm stained with Hoechst 33342 prior to insemination is observed on the surface
of the inner perivitelline layer (PL). The sperm's acrosome will release a trypsin like enzyme, acrosin, and digest a hole
through the inner PL. The right panel shows multiple sperm holes (white perforations) in the inner PL overlying the
germinal disc (GD) of a duck ovum (polyspermy is normal in birds). Sperm hole numbers can be used to assess true
fertility and the duration of the fertile period.
Unlike mammals, polyspermy is the norm in avian fertilization. The GD (3.5 mm in diameter)
provides a relatively small target for fertilization in the large megalecithal ova (yolk-filled ova)
of chickens and turkeys (3.5 – 4.0 mm in diameter); thus polyspermy may be an evolutionary
adaptation to ensure higher rates of fertilization in such species [74]. The inner PL may be
penetrated by many sperm, although only one male pronucleus will ultimately fuse (syngamy)
with the female pronucleus to form the nascent embryo (reviewed in [75-77]. A single sperm
hole in the inner PL does not ensure fertilization. Although turkeys show a lower number of
sperm interacting with ova relative to chickens, the presence of three sperm holes in the inner
PL predicts a 50% probability of fertilization, whereas, six sperm holes suggest a probability
greater than 95% fertilization [78]. The outer PL is rapidly depositied around the ovum in the
posterior infundibulum and proximal magnum and is impenetrable by sperm [21, 78-79] thus
preventing pathological polyspermy.
Given the volume of the GD relative to a single sperm, another possible function of polyspermy
may be to activate specific molecular factors in the GD cytoplasm thereby initiating the process
of embryogenesis. Yet, polyspermy also results in the presence of multiple male pronuclei in
the GD. To cope with this potentially harmful scenario, the mature ovum has been found to
have DNase I and II endonuclease activities, both of which will degrade sperm DNA [76]. In
contrast, no such DNase activity has been detected in mammalian ova that engage in mono‐
spermic fertilization, further suggesting the role of these enzymes in the avian embryo is to
protect against detrimental genetic consequences of polyspermy [76].
The number of holes in the inner PL is highly positively correlated with fertility. Correlations
exist between the number of sperm inseminated, the number undergoing the acrosome
reaction at the inner PL [80], and the number of sperm embedded in the outer PL [81]. The
number of sperm holes in the inner PL and the number sperm trapped in the outer PL may be
used to estimate the duration of fertility (‘fertile period') in hens. While the number of sperm
penetrating the inner PL shows a decreasing logarithmic relationship over time [81-82], a
positive correlation between the total number of sperm penetrating the inner PL and the
number of sperm stored in the SSTs was observed [83]. Given these observations, it should not
be surprising there is also a positive correlation between the number of SSTs containing sperm
and the proportion of sperm that have undergone the acrosome reaction at the inner PL [82].
Success in Artificial Insemination - Quality of Semen and Diagnostics Employed
3. Techniques in artificial insemination and fertility evaluation in poultry
For non-domestic birds, chapters in Bakst and Long [4], Lake and Stewart [5] and Bakst and
Wishart [6] provide overviews of semen evaluation and AI techniques. Artificial insemination
technology and reproductive biology for ratites were reviewed by Malecki et al. [84].
3.1. Semen collection
Primarily due to the anatomical variation of the phallic region in different birds, semen
collection techniques will vary. In contrast to ratites and water-fowl with an intromittent
phallus, Galliformes (chicken, turkey, and quail) do not have an intermittent organ. Their non-
intromittent organ consists of folds and bulges that make contact with the female's cloaca at
mating. From an anatomical perspective, there are considerable differences between the non-
intromittent organs of the chicken and turkey (Figure 6). The rooster has a prominent medial
phallic body and relatively small lateral phallic bodies and lymph folds. Conversely, the turkey
tom has no medial phallic body but prominent lateral phallic bodies and lymph folds. Sex
sorting at hatch by cloacal examination is based on the relative differences in size of these
structures between the males and females.
Figure 6. The turkey (left) and chicken (right) cloacae are viewed with the dorsal lips of the cloacae pulled back to
expose each species phallus non-protrudens. Unlike the turkey, the chicken has a central protuberance, the medial
phallic body (MPB) and regressed lateral phallic bodies (LPB) and lymph folds (LF). The turkey phallus non-protrudens
is characterized by dominant LPB and LF and the conspicuous absence of the MPB.
The goal of semen collector is to obtain the maximum volume of clean, high quality semen
with the minimal amount of handling. In chickens and turkeys, the abdominal massage
technique [1, 4] involves massaging the cloacal region to achieve phallic tumescence. This is
followed by a ‘cloacal stroke', a squeezing of the region surrounding the sides of the cloaca to
express the semen. Little additional semen can be expressed after two cloacal strokes; addi‐
tional cloacal strokes may cause damage to the phallic and cloacal regions and contribute to
semen contamination [85].
Artificial Insemination in Poultry
Semen should be pearly white, viscous, and clean. With each male collected, the semen collector
should perform a visual examination of the semen at the time of ejaculation. This is easier with
the turkey because the ejaculate accumulates on the phallus before it is collected by the ‘milker'
(semen collector). Off-color or watery semen, and semen contaminated with blood or fecal/
urates debris should not be used for insemination. Due to the increased volume of transparent
fluid in rooster semen, which is a transudate derived from the phallus at the time of ejaculation,
chicken semen is less viscous and sperm concentration lower than that of turkey semen.
3.2. Sperm concentration
If semen is to be diluted, it is best .to have a known volume of semen diluent (a tissue culture-
like medium formulated to sustain sperm viability) at ambient temperature in the semen
receptacle before collection begins. For routine AI of turkey hens, semen from 10-12 toms are
pooled in a single receptacle, mixing the semen gently after each male is collected. Semen
volume is determined and if the AI dose is based on numbers of sperm (generally 250-350
million sperm per dose) sperm concentration is determined. The most popular techniques for
determining sperm concentration are the packed cell volume (PCV; also referred to as a
spermatocrit) and optical density (OD; photometry).
Determining sperm concentration using PCVs is nearly identical to that of determining blood
hematocrit values. Semen aspirated into micro-hematocrit tubes are centrifuged in a hema‐
tocrit centrifuge until the sperm are tightly packed (10 min); the percentage of packed sperm
cells relative to the original semen volume in the micro-tube is determined. Sperm concentra‐
tion is derived using a conversion factor or standard curve previously derived by comparing
and graphically plotting varying ascending sperm concentrations from hemocytometer counts
to corresponding spermatocrit readings. (See [4] for detailed protocols to determine sperm
concentration and the derivation of standard curves.)The optical density (OD) is determined using a photometer. The OD of highly diluted semen
is directly proportional to the concentration of sperm, thus providing an indirect estimate of
the sperm concentration. Like the PCV method, sperm concentration is derived using a
conversion factor or previously derived standard curve by comparing and graphically plotting
varying sperm concentrations from hemocytometer counts to corresponding OD readings [4].
The PVC and OD methods are two indirect methods of determining sperm concentration, that
is, the final concentration is calculated from a regression equation or standard curve derived,
in part, from direct sperm counts with a hemocytometer [4]. Briefly, to derive a regression
equation and standard curve, serial dilutions (n=5) covering a wide range of sperm concen‐
trations are prepared and sperm concentrations are determined with a hemocytometer and
the instrument or method that requires the standard curve (at least 4 replicates with 4 different
semen samples). This is a tedious procedure but if reliable and repeatable sperm numbers are
to be inseminated it is best to establish standard curves for each instrument every 12-18 months.
The reason for this is that the rotational speed of different centrifuges and the intensity of a
photometer's light source may differ as a result of manufacturer's variation, age of the
instrument, and/or repeated use of the instrument, thereby producing variations in the
respective final readings and subsequent calculations of sperm concentrations.
Success in Artificial Insemination - Quality of Semen and Diagnostics Employed
Another concern when using any semen evaluation method is variation in the operator's
techniques. Consistency is the key to repeatable data. The technical staff all must follow the
same standard operating procedures (SOPs). For example, when counting sperm with a
hemocytometer, all individuals in a lab should following the same SOP for how long the sperm
are permitted to settle on the grid and which sperm to count or omit from the count. Also, is
the photometer zeroed with the same buffer? If a procedure calls for an incubation period,
such as in a live-dead stain, are the samples being incubated for the same duration each time
using the same stain concentrations? A lack of consistency in following the SOPs within a
laboratory will lead to unwarranted variation and non-reproducible and inaccurate data.
3.3. Sperm viability
In the context of semen evaluation, reference to ‘viable' sperm simply implies that such
sperm possess an intact plasmalemma and are assumed to be functional. Plasmalemma in‐
tegrity is frequently determined using either a dead-cell or a live-cell stain alone or simulta‐
neously. The dead-cell stains are excluded by sperm with an intact plasmalemma but stain
dead sperm possessing a permeable plasmalemma. Live-cell stains permeate the intact
sperm plasmalemma and become visible only after reacting with cytosolic enzymes or inter‐
acting with sperm nuclear proteins. Both eosin and propidium iodide are popular dead-cell
stains while calcein AM and SYBR-14 are frequently used live-cell stains (see [86] for extend‐
ed discussion and availability for the live-cell probes). On a commercial breeder farm, the
nigrosin/eosin (N/E) technique is most likely the procedure to be used to determine sperm
viability [4]. Briefly, sperm are stained with N/E and a smear of the stained sperm is made
on a slide (Figure 7). Under a bright field microscope the viable sperm remain pearly white,
while eosin will stain non-viable sperm a pink to magenta color. The nigrosin serves as a
background to enhance differentiation between the non-viable and viable sperm. In contrast
to the N/E technique, a more sophisticated laboratory may use flow cytometry that sorts via‐
ble from non-viable sperm after staining with calcein AM or SYBR-14 and propidium iodide.
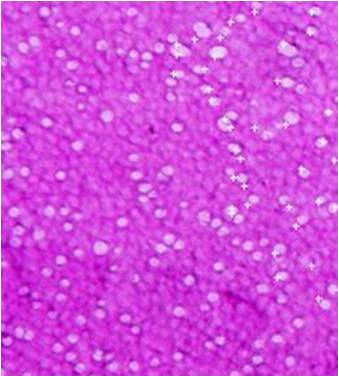
Figure 7. The left panel shows a nigrosin eosin preparation of turkey sperm with nearly 100% viable sperm (un‐
stained) white nuclei and midpieces. The sperm head is clearly visible as the white arcing segment; the acrosome and
midpiece are difficult to differentiate from the nucleus. The upper right panel reveals a normal sperm and a second
sperm with an abnormally curved and swollen midpiece. Observed in the lower right panel is a nonviable sperm
stained with eosin throughout the nucleus and midpiece. Barely visible at the anterior end of the nucleus is the un‐
stained, conical shaped acrosome.
Artificial Insemination in Poultry
3.4. Sperm motility and mobility
Sperm motility can be progressive (forward direction) or non-progressive (random movement
or oscillations) movement. Generally, progressive motility is determined subjectively at
ambient temperature using a microscope at low magnification (hanging-drop technique) or
objectively using a computer-assisted semen analysis system. These techniques are reviewed
by Bakst and Long [4]. Motility evaluated by microscopy has been shown to have little
correlation with fertility and simply reveals that the sperm are motile. First described by
Froman and McLean [87] and further elaborated for commercial use by Froman [88], the sperm
mobility assay has gained popularity as a measure of an individual male's ability to produce
highly mobile sperm [mobility defines the ability of sperm to move progressively against a
viscous medium (Accudenz) at 41°C] that are more likely to fertilize an ovum than males
producing less mobile sperm. While the sperm mobility assay is a powerful tool for the
selection of the most fecund males to be used in AI, it necessitates attention to details and
accurate and consistent preparation of the reagents.
3.5. Evaluation of fertility
The measure of a successful AI program is sustained hen fertility. While candling-fertility is
useful, there is an eight or more day lag between the last AI and candling-fertility determina‐
tion, which overlaps with the next insemination (hen insemination is generally at 7-day
intervals). With AI programs, it is often desirable to determine the fertility status of a flock
before the next weekly insemination. There are several options available: breaking-out fresh
eggs and examining the GD to differentiate a fertilized from an unfertilized or early dead
embryo; setting normal but culled eggs (checked, hairline cracked, or dirty eggs) in a spare
incubator for 24-36 hr before breaking-out [89]; counting sperm in the outer PL; and counting
sperm holes in the inner PL. The above procedures are reviewed in Bakst and Long [4].
As noted previously, the sperm penetration assay is not only used to determine true fertility,
but also to estimate the number of sperm residing in the use SSTs at the time of ovulation [90].
The isolation of the inner PL and staining procedure, initially developed for chicken eggs by
Bramwell et al. [91], was quickly adapted to turkey eggs by Donoghue [92]. The major
drawback to the sperm penetration assay as originally described is that it is time consuming,
particularly with respect to isolating, washing, and positioning the PL wrinkle-free on the slide.
Spasojevic [10] and colleagues at Willmar Poultry Company (Willmar, MN) significantly
increased the efficiency of preparing the PL slides from turkey eggs in the following manner:
the albumen is removed from the ovum as in the original procedure [4]; a square is outlined
on a slide using super glue; the slide is placed firmly on the ovum's surface with the GD
centered in the square; after the glue is set, the PL is cut and washed to remove adhering yolk.
The advantages here are speed and the PL remains wrinkle free.
A different modification of the sperm penetration assay was suggested by I.A. Malecki
(personal communication) and entails placing a filter ring over the GD (inside diameter slightly
larger than the GD), cutting around the outside diameter of the filter ring (about 2 mm between
the inside and outside perimeter of the ring), and lifting the filter ring off the ovum. The filter
ring with the adhering PL is washed gently with saline to remove the yolk and GD material
Success in Artificial Insemination - Quality of Semen and Diagnostics Employed
until transparent, placed on a slide, and then fixed and stained with saline washes after each
step. Our laboratory has used the filter ring technique with eggs from broilers, turkeys, ducks,
and quail and it is now our preferred method for the performing the sperm penetration assay.
Artificial insemination is a common practice in the poultry industry with the turkey industry
in North America and Europe using it almost exclusively for the production of hatching eggs.
The broiler industry has not adapted AI for several reasons: because of sheer numbers of broiler
breeders that need to be inseminated weekly, the labor cost would be very significantly; the
initial investment in special housing for the males; an efficient, cost effective means of actually
performing the inseminations (housing and catching the hens) would need to be developed;
and finally, the concern that after a few generations of breeding broilers by AI, the behaviors
associated with natural mating may be less dominant. Notwithstanding these concerns, the
benefits of AI for broilers would include the following: the male:female ratio would be increase
from 1:10 for natural mating to 1:25 with AI; with fewer males needed, there would be greater
selection pressure on the male traits of economic importance and subsequently greater genetic
advancement per generation; biosecurity concerns associated with "spiking" aging hen flocks
with new and/or younger males to augment mating frequency and fertility would be elimi‐
nated; and, differences in body conformation between males and females that impact semen
transfer at mating would no longer be a consideration.
In 1995, Sir Peter Lake wrote an excellent review of the history of AI, its impact on the poultry
industry, and what is needed to advance the practice of AI with poultry [93]. Unfortunately,
AI technology has not advanced significant since this review article. More than 15 years later,
the only significant advance is in the evaluation of sperm mobility and the impact that males
producing highly mobile sperm have on paternity [9]. Notwithstanding, it is foreseeable that
sometime in the future, research addressing poultry sperm biology and the cellular and
molecular basis of oviductal sperm transport, selection, and storage will lead to the following
innovations in AI technology: insemination intervals increased to 10-14 days (versus 7-day)
with fewer sperm per insemination; in vitro sperm storage for 24-36 hr at ambient temperature
with minimal loss of sperm viability; and, the possibility of transgenic progeny following the
insemination of sperm carrying transgenes.
M.R. Bakst* and J.S. Dymond
*Address all correspondence to: [email protected]
Animal Biosciences and Biotechnology Laboratory Beltsville Area, Agricultural Research
Service, U.S. Department of Agriculture Beltsville, Maryland, USA
Artificial Insemination in Poultry
[1] Burrows WA, Quinn JP. The method of obtaining spermatoza from the domestic
fowl. Poultry Science 1935;14(4) 251-4.
[2] Quinn JP, Burrows WA. Artificial insemination in fowls. The Journal of Heredity
1936;27(1) 31-7.
[3] Spasojevic R. Two hundred million sperm cells per hen? No Way! Proceedings – Mid‐
west Poultry Federation Convention, Turkey Breeder Workshop, March 16-18, 2010. CD
available at MPF. St. Paul, Minnesota. www.midwestpoultry.com
[4] Bakst MR, Long JA. Techniques for Semen Evaluation, Semen Storage, and Fertility
Determination. 2, 1-113. 2010. St. Paul, Minnesota, The Midwest Poultry Federation.
[5] Lake PE, Stewart JM. Artificial insemination in poultry. Bulletin 213. 1978. London,
UK, Ministry of Agriculture, Fish, and Food. Her Majesty's Stationary Office.
[6] Bakst MR, Wishart GJ. Proceedings, First Internation Symposium on the Artificial In‐
semination of Poultry. 1, 1-297. 1995. Champaign, IL, Poultry Science Association,
[7] Bakst MR, Cecil H. Techniques for Semen Evaluation, Semen Storage, and Fertility
Determination. 1-97. 1997. Savoy, IL, Poultry Science Association.
[8] Jamieson BGM. Reproductive Biology and Phylogeny of Birds. Enfield, New Hamp‐
shire: Science Publishers; 2007.
[9] Froman DP, Feltmann AJ, Pendarvis K, Cooksey AM, Burgess SC, Rhoads DD. A
proteome-based model for sperm mobility phenotype. Journal of Animal Science
2011;89(5) 1330-7.
[10] Bakst MR. Physiology and Endocrinology Symposium: Role of the oviduct in main‐
taining sustained fertility in hens. Journal of Animal Science 2011;89(5)1323-9.
[11] Perry MM, Waddington D, Gilbert AB, Hardie MA. Growth rates of the small yolky
follicles in the ovary of the domestic fowl. IRCS Medical Science: Experimental Ani‐
mals 1983;11(11/12) 979-80.
[12] Saito N, Sato K, Shimada K. Prostaglandin levels in peripheral and follicular plasma,
the isolated theca and granulosa layers of pre- and postovulatory follicles, and the
myometrium and mucosa of the shell gland (uterus) during a midsequence-oviposi‐
tion of the hen (Gallus domesticus). Biology of Reproduction 1987;36(1) 89-96.
[13] Dick HR, Culbert J, Wells JW, Gilbert AB, Davidson MF. Steroid hormones in the
postovulatory follicle of the domestic fowl (Gallus domesticus). Journal of Reproduc‐
tion and Fertility 1978;53(1) 103-8.
[14] Hammond RW, Olson DM, Frenkel RB, Biellier HV, Hertenlendy F. Prostaglandins
and steroid hormones in plasma and ovarian follicles during the ovulation cycle of
Success in Artificial Insemination - Quality of Semen and Diagnostics Employed
the domestic hen (Gallus domesticus). General and Comparative Endocrinology
1980;42(2) 195-202.
[15] Hertenlendy F, Biellier HV, Todd H. Effects of the egg cycle and the route of admin‐
istration on prostaglandin-induced oviposition of hens and Japanese quail. Journal of
Reproduction and Fertility 1975;44(3) 579-82.
[16] Johnson AL, Woods DC. Ovarian dynamics and follicle development. In: Jamieson
BGM, editor. Reproductive Biology and Phylogeny of Birds.Enfield, New Hamp‐
shire: Science Publishers; 2007. p. 243-77.
[17] Wyburn GM, Johnston HS, Draper MH, Davidson MF. The fine structure of the in‐
fundibulum and magnum of the oviduct of Gallus domesticus. Quarterly Journal of
Experimental Physiology 1970;55(3):213-32.
[18] Perry MM. Nuclear events from fertilisation to the early cleavage stages in the do‐
mestic fowl (Gallus domesticus). Journal of Anatomy 1987;150(1) 99-109.
[19] Birkhead TR, Sheldon BC, Fletcher F. A comparative study of sperm–egg interactions
in birds. Journal of Reproduction and Fertility 1994;101(2) 353-61.
[20] Bakst MR, Howarth B, Jr. Hydrolysis of the hen's perivitelline layer by cock sperm in
vitro. Biology of Reproduction 1977;17(3) 370-9.
[21] Okamura F, Nishiyama H. The passage of spermatozoa through the vitelline mem‐
brane in the domestic fowl, Gallus gallus. Cell and Tissue Research 1978;188(3)
[22] Bakst MR, Wishart G, Brillard JP. Oviductal sperm selection, transport, and storage
in poultry. Poultry Science Reviews 1994;5 117-43.
[23] Froman DP, Feltmann AJ, McLean DJ. Increased fecundity resulting from semen do‐
nor selection based upon in vitro sperm motility. Poultry Science 1997;77(1) 73-7.
[24] Bakst MR. Observations on the turkey oviductal sperm-storage tubule using differen‐
tial interference contrast microscopy. Journal of Reproduction and Fertility 1992;95(3)
[25] Schuppin GT, Van Krey HP, Denbow DM, Bakst MR, Meyer GB. Ultrastructural
analyses of uterovaginal sperm storage glands in fertile and infertile turkey breeder
hens. Poultry Science 1984;63(9) 1872-82.
[26] Bakst MR, Akuffo V. Alkaline phosphatase reactivity in the vagina and uterovaginal
junction sperm-storage tubules of turkeys in egg production: Implication for sperm
storage. British Poultry Science 2007;48(4) 515-8.
[27] Brillard JP. Sperm storage and transport following natural mating and artificial in‐
semination. Poultry Science 1993;72(5) 923-8.
Artificial Insemination in Poultry
[28] Christensen VL, Bagley LG. Efficacy of fertilization in artificially inseminated turkey
hens. Poultry Science 1989;68(5) 724-9.
[29] Bakst MR, Donoghue AM, Yoho DE, Moyle JR, Whipple SM, Camp MJ, et al. Com‐
parisons of sperm storage tubule distribution and number in 4 strains of mature
broiler breeders and in turkey hens before and after the onset of photostimulation.
Poultry Science 2010;89(5) 986-92.
[30] Birkhead TR, Moller AP. Numbers and size of sperm storage tubules and the dura‐
tion of sperm storage in birds: A comparative study. Biological Journal of the Lin‐
nean Society 1992;45(4):363-72.
[31] Brillard JP, Beaumont C, Scheller MF. Physiological responses of hens divergently se‐
lected on the number of chicks obtained from a single insemination. Journal of Re‐
production and Fertility 1998;114(1) 111-7.
[32] Bakst MR. Oviducal sperm selection, transport, and storage in poultry: A review. Re‐
production Fertility and Development 1993;5(6) 595-9.
[33] Blesbois E, Brillard JP. Specific features of in vivo and in vitro sperm storage in birds.
[34] Van Krey HP, Ogasawara FX, Pangborn J. Light and electron microscopic studies of
possible sperm gland emptying mechanisms. Poultry Science 1967;46(1) 69-78.
[35] Fujii S. Histological and histochemical studies on the oviduct of the domestic fowl
with special reference to the region of utero-vaginal junction. Archives of Histology
and Cytology 1963;23(5) 447-9.
[36] Renden JA, May EB, Benoff FH. Histochemistry of uterovaginal sperm-host glands in
Japanese quail (Coturnix coturnix japonica) with reference to the period of oviposition.
Poultry Sci 1981;60(11) 2529-35.
[37] Blesbois E, Lessire M, Grasseau J, Hallouis JM, Hermier D. Effect of dietary fat on the
fatty acid composition and fertilizing ability of fowl semen. Biology of Reproduction
1997;56(5) 1216-20.
[38] Surai PF. Natural Antioxidants in Avian Nutrition and Reproduction. Nottingham,
UK: Nottingham University Press; 2002.
[39] Williams K, Frayne J, Hall L. Expression of extracellular glutathione peroxidase type
5 (GPX5) in the rat male reproductive tract. Molecular Human Reproduction
1998;4(9) 841-8.
[40] Catalá A. Lipid peroxidation modifies the picture of membranes from the "Fluid Mo‐
saic Model" to the "lipid whisker model". Biochimie 2012;94(1) 101-9.
[41] Breque C, Surai P, Brillard JP. Roles of antioxidants on prolonged storage of avian
spermatoza in vivo and in vitro. Molecular Reproduction and Development 2003;66(3)
Success in Artificial Insemination - Quality of Semen and Diagnostics Employed
[42] Long C, Sonstegard TS, Long JA, van Tassell CP, Zuelke KA. Serial analysis of gene
expression in turkey sperm storage tubules in the presence and absence of resident
sperm. Biology of Reproduction 2003;69(2) 469-74.
[43] Foye-Jackson OT, Long JA, Bakst MR, Blomberg LA, Akuffo V, Silva VB, et al. Ovi‐
ductal expression of avidin, avidin-related protein-2, and progesterone receptor in
turkey hens in relation to sperm storage: Effects of oviduct tissue type, sperm pres‐
ence, and turkey line. Poultry Science 2011;90(7) 1539-47.
[44] O'Malley BW, McGuire WL. Studies on the mechanism of action of progesterone in
regulation of the synthesis of specific protein. Journal of Clinical Investigation
1968;47(3) 654-64.
[45] Korpela JK, Elo HA, Tuohimaa PJ. Aviadin induction by estrogen and progesterone
in te immature oviduct of chicken, Japanese quail, duck, and gull. General and Com‐
parative Endocrinology 1981;44(2) 230-2.
[46] Burke WH, Ogasawara FX. Presence of spermatozoa in uterovaginal fluids of the hen
at various stages of the ovulatory cycle. Poultry Science 1969;48(2) 408-13.
[47] Compton MM, Van Krey HP, Siegel PB. The filling and emptying of the uterovaginal
sperm-host glands in the domestic hen. Poultry Science 1978;57(6) 1696-700.
[48] Bushman AF, Van Krey HP, Denbow DM, Siegel PB. Effect of the ovulatory cycle on
oviductal sperm storage in the domestic fowl. Theirogenology 1985;23(3) 473-9.
[49] Bakst MR. Duration of fertility of turkeys inseminated at different times after the on‐
set of photostimulation. Journal of Reproduction and Fertility 1988;84(2) 531-7.
[50] Das SC, Nagasaka N, Yoshimura Y. Changes in the expression of estrogen receptor
mRNA in the utero-vaginal junction containing sperm storage tubules in laying hens
after repeated artifical insemination. Theriogenology 2006;64(4) 893-900.
[51] Yoshimura Y, Koike K, Okamoto T Immunolocalization of progesterone and estro‐
gen receptors in the sperm storage tubules of laying and diethylstilbestrol-injected
immature hens. Poultry Science 2000;79(1) 94-8.
[52] Ito T, Yoshizaki N, Tokumoto T, Ono H, Yoshimura Y, Tsukada A, et al. Progester‐
one is a sperm-releasing factor from the sperm-storage tubules in birds. Endocrinolo‐
gy 2011;152(10) 3952-62.
[53] Agre P, Sasaki S, Chrispeels MJ. Aquaporins: A family of water channel proteins.
American Journal of Physiology Renal Physiology 1993;265(3) F461.
[54] Zaniboni L, Bakst MR. Localization of aquaporins in the sperm storage tubules in the
turkey oviduct. Poultry Science 2004;83(7) 1209-12.
[55] Froman DP, Bowling ER, Wilson JL. Sperm mobility phenotype not determined by
sperm quality index. Poultry Science 2003;82(3) 496-502.
Artificial Insemination in Poultry
[56] Froman DP. Effect of sperm mobility phenotype on fertility, sperm competition, and
in vivo sperm storage in the domestic fowl. Journal of Dairy Science 93[Suppl. 1],
[57] Brillard JP, Galut O, Nys Y. Possible causes of subfertility in hens following insemi‐
nation near the time of oviposition. British Poultry Science 1987;28(2) 307-18.
[58] Stephens RE, Prior G. Dynein from serotonin-activated cilla and flagella: Extraction
characteristics and distinct sites for cAMP-dependent protein phosphorylation. Jour‐
nal of Cell Science 1992;103(4):999-1012.
[59] Holm L, Ridderstrale Y, Knutsson PG. Localisation of carbonic anhydrase in the
sperm storing regions of the domestic hen. Cell Tissues Organs 1996;156(4):253-60.
[60] Holm L, Wishart GJ. The effect of pH on the motility of spermatoza from chicken,
turkey and quail. Animal Reproduction Science 1998;54(1):45-54.
[61] Freedman SL, Akuffo VG, Bakst MR. Evidence for the innervations of sperm storage
tubules in the oviduct of the turkey (Meleagris gallopavo). Reproduction 2001;121(5):
[62] Bakst MR, Akuffo V. Serotonin localization in the turkey vaginal but not sperm stor‐
age tubule epithelia. Journal of Avian Biology 2008;39(3):348-54.
[63] Froman DP, Bowling ER, Wilson JL. Sperm mobility phenotype not determined by
sperm quality index. Poultry Science 2003;82(3):496-502.
[64] Froman DP, Engel HN. Alteration of the spermatozoal glycocalyx and its effect on
duration of fertility in the fowl (Gallus domesticus). Biology of Reproduction
[65] Froman DP, Thursam KA. Desialylation of the rooster sperm's glycocalyx decreases
sperm sequestration following intravaginal insemination of the hen. Biology of Re‐
[66] Steele MG, Wishart GJ. Demonstration that the removal of sialic acid from the sur‐
face of chicken spermatozoa impedes their transvaginal migration. Theriogenology
[67] Pelaez J, Long JA. Characterizing the glycocalyx of poultry spermatozoa: I. Identifica‐
tion and distribution of carbohydrate residues using flow cytometry and epifluores‐
cence microscopy. Journal of Andrology 2007;28(2):342-52.
[68] Robertson L, Wishart GJ, Horrocks AJ. Identification of perivitelline N-linked gly‐
cans as mediators of sperm-egg interaction in chickens. Journal of Reproduction and
[69] Birkhead TR, Brillard JP. Reproductive isolation in birds: postcopulatory prezygotic
barriers. Trends in Ecology & Evolution 2007;22(5):266-72.
Success in Artificial Insemination - Quality of Semen and Diagnostics Employed
[70] Steele MG, Meldrum W, Brillard JP, Wishart GJ. The interaction of avian spermato‐
zoa with the perivitelline layer in vitro and in vivo. Journal of Reproduction and Fer‐
[71] Howarth B. Carbohydrate involvement in sperm-egg interaction in the chicken. Jour‐
nal of Receptor Research 1992;12(2):255-65.
[72] Horrocks AJ, Stewart JM, Jackson L, Wishart GJ. Induction of acrosomal exocytosis in
chicken spermatozoa by inner perivitelline-derived N-linked glycans. Biochemical
and Biophysical Research Communications 2000;278(1):84-9.
[73] Koyanagi F, Masuda S, Nishiyama H. Acrosome reaction to cock spermatoza incu‐
bated with the perivitelline layer of the hen's ovum. Poultry Science 1988;67(12):
[74] Bakst MR, Howarth B, Jr. The fine structure of the hen's ovum at ovulation. Biology
of Reproduction 1977;17(3):361-9.
[75] Wasserman PM, Jovine L, Litscher ES. A profile of fertilization in mammals. Nature
Cell Biology 2001;3(2):E59-64.
[76] Stepinska U, Olszanska B. DNase I and II present in avian oocytes: A possible in‐
volvement in sperm degradation at polyspermic fertilisation. Zygote 2003;11(1):
[77] Stepinska U, Bakst MR. Fertilization. In: Jamieson BGM, editor. Reproductive Biolo‐
gy and Phylogeny of Birds. Enfield, New Hampshire: Science Publishers; 2007. p.
[78] Wishart GJ. Quantitative aspects of sperm:egg interaction in chickens and turkeys.
Animal Reproduction Science 1997;48(1):81-92.
[79] Bellaris R, Harkness M, Harkness RD. The vitelline membrane of the hen's egg: A
chemical and electron microscopal study. Journal of Ultrastructure Research
[80] Bramwell RK, Howarth B, Jr. Preferential attachment of cock spermatozoa to the per‐
ivitelline layer directly over the germinal disc of the hen's ovum. Biology of Repro‐
[81] Wishart GJ. Regulation of the length of the fertile period in the domestic fowl by
numbers of oviducal spermatozoa, as reflected by those trapped in laid eggs. Journal
of Reproduction and Fertility 1987;81(2):495-9.
[82] Brillard JP, Antoine H. Storage of sperm in the uterovaginal junction and its inci‐
dence on the numbers of spermatoza present in the perivitelline layer of hen's eggs.
British Poultry Science 1990;31(3):635-44.
[83] Wishart GJ. Quantitative aspects of sperm:egg interaction in chickens and turkeys.
Animal Reproduction Science 1997;48(1):81-92.
Artificial Insemination in Poultry
[84] Malecki IA, Rybnik PK, Martin GB, Malecki I, Glatz P. Artificial insemination tech‐
nology for ratites: A review. Australian Journal of Experimental Biology 2008;48(10):
[85] Bakst MR, Cecil HC. Gross appearance of turkey cloacae before and after single or
multiple manual semen collections. Poultry Science 1983;62(4):683-9.
[86] Haugland RP. Handbook of Fluorescent Probes and Research Products. 9th ed. Eu‐
gene, Oregon: Molecular Probes, Inc.; 2002.
[87] Froman DP, McLean DJ. Objective measurement of sperm motility based upon
sperm penetration of Accudenz®. Poultry Science 1996;75(6):776-84.
[88] Froman DP. Application of the sperm mobility assay to primary broiler breeder
stock. Journal of Applied Poultry Research 2006;15(2):280-6.
[89] Bakst MR, McGary S, Estevez I, Knapp T. Use of nonsettable eggs to evaluate turkey
hen fertility. Journal of Applied Poultry Research 2002;11(4):402-5.
[90] Brillard JP, Bakst MR. Quantification of spermatozoa in the sperm-storage tubules of
turkey hens and the relation to sperm numbers in the perivitelline layer of eggs. Biol‐
ogy of Reproduction 1990;43(2):271-5.
[91] Bramwell RK, Marks HL, Howarth B. Quantitative determination of spermatozoa
penetration of the perivitelline layer of the hen's ovum as assessed on oviposited
eggs. Poultry Science 1995;74(11):1875-83.
[92] Donoghue AM. The effect of twenty-four hour in vitro storage on sperm hydrolysis
through the perivitelline layer of ovipositioned turkey eggs. Poultry Science
[93] Lake, PE. Historical perspective of artificial insemination technology. In: Bakst MR
and Wishart GJ, editors. Proceedings - First International Symposium on the Artifical
Insemination of Poultry. Savoy, Illinois: Poultry Science Association; 1995. p. 1-20
Source: http://www.vetrade.ir/files/56634/gallery_file/file_1455275607.pdf
Advances in Interventions for Families with a Relative with a Personality Disorder Diagnosis Perry D. Hoffman, PhD, and Alan E. Fruzzetti, PhD and Harvey [4] found that these family members report Perry D. Hoffman, PhDPresident, NEA-BPD, 734 East Boston Post Road, Mamaroneck, higher levels of psychological distress compared with community norms. However, the challenges facing family
Number 18 n July 30, 2009 Costs of Complementary and Alternative Medicine (CAM) and Frequency of Visits to CAM Practitioners: United States, 2007 by Richard L. Nahin, Ph.D., M.P.H., National Institutes of Health; Patricia M. Barnes, M.A.;