Kamagra gibt es auch als Kautabletten, die sich schneller auflösen als normale Pillen. Manche Patienten empfinden das als angenehmer. Wer sich informieren will, findet Hinweise unter kamagra kautabletten.
Pq089603275p
Proc. Natl. Acad. Sci. USA
Vol. 93, pp. 3275–3279, April 1996
DUB-1, a deubiquitinating enzyme with growth-
suppressing activity
YUAN ZHU*, MARTIN CARROLL*, FEROZ R. PAPA†, MARK HOCHSTRASSER†, AND ALAN D. D'ANDREA*‡*Divisions of Pediatric Oncology and Cellular and Molecular Biology, Dana–Farber Cancer Institute, Harvard Medical School, Boston, MA 02115; and
†Department of Biochemistry and Molecular Biology, The University of Chicago, Chicago, IL 60637
Communicated by David M. Livingston, Dana–Farber Cancer Institute, Boston, MA, November 29, 1995 (received for review October 3, 1995)
Cytokines regulate cell growth by inducing
substrate. These enzymes are ubiquitin specific but share
the expression of specific target genes. Using the differential
certain properties with other thiol proteases. Genes for at least
display method, we have cloned a cytokine-inducible imme-
15 deubiquitinating enzymes were identified from the yeast
diate early gene, DUB-1 (for deubiquitinating enzyme).
genome, making them the largest known gene family in the
DUB-1 is related to members of the UBP superfamily of
ubiquitin system. Several proteins implicated in growth and
deubiquitinating enzymes, which includes the oncoprotein
development, including the mammalian proteins Tre-2 and
Tre-2. A glutathione S-transferase–DUB-1 fusion protein
Unp and the
Drosophila fat facets protein, were either shown
cleaved ubiquitin from a ubiquitin-b
-galactosidase protein.
to be deubiquitinating enzymes or to have sequence similarity
When a conserved cysteine residue of DUB-1, required for
to such enzymes (21).
ubiquitin-specific thiol protease activity, was mutated to
In the current study, we used the strategy of differential
serine (C60S), deubiquitinating activity was abolished. Con-
display (22, 23) to clone an immediate early cDNA (
DUB-1)
tinuous expression of DUB-1 from a steroid-inducible pro-
that is specifically induced by IL-3. The
DUB-1 cDNA encodes
moter induced growth arrest in the G1 phase of the cell cycle.
a 526-aa protein that has deubiquitinating activity. Interest-
Cells arrested by DUB-1 expression remained viable and
ingly, misregulated expression of
DUB-1 induces cell cycle
resumed proliferation upon steroid withdrawal. Our results
suggest that DUB-1 regulates cellular growth by modulating
arrest in the G1 phase of the cell cycle. Our results support the
either the ubiquitin-dependent proteolysis or the ubiquitina-
hypothesis that protein ubiquitination is important in growth-
tion state of an unknown growth regulatory factor(s).
factor–mediated cellular proliferation. They also implicate
deubiquitinating enzymes as regulatory enzymes that couple
Interleukin 3 (IL-3) is a glycoprotein hormone that regulates
extracellular signaling to cell growth.
growth of hematopoietic progenitor cells (1). IL-3, like other
cytokines, acts during the G1 phase of the cell cycle to drive
MATERIALS AND METHODS
cells into S phase. IL-3 exerts its biologic function through a
specific receptor (IL-3R) that is expressed on its target cells (2,
Cells and Cell Culture. BayF3 is an IL-3-dependent murine
3). The IL-3R activates multiple signal transduction pathways,
pro-B cell line (24). BayF3 cells were maintained in RPMI
including the Ras–Raf–mitogen activating protein kinase path-
1640 medium supplemented with 10% fetal calf serum (FCS)
way and the JAK–STAT pathway, resulting in the induction of
and 10% conditioned medium from WEHI-3B cells as a source
immediate early genes. How these immediate early genes
of IL-3 (25).
couple IL-3R activation to the biochemical machinery of cell
Differential Display and Cloning of DUB-1 cDNA. Total
growth and cell cycle progression is poorly understood.
cellular RNA was isolated from starved or IL-3-stimulated
Cell growth and cell cycle progression are controlled, at least
BayF3 cells by the guanidinium isothiocyanate procedure (26)
in part, by ubiquitin-mediated proteolysis (4, 5). Ubiquitin-
and subjected to the differential display analysis (22) (Gene
mediated proteolysis requires ATP and results in covalent
Hunter, Boston). A partial cDNA fragment that was specifi-
conjugation of target proteins with multiple ubiquitin mole-
cally induced by IL-3 was isolated using a 59 primer (59-
cules (6–9). Multiubiquitinated proteins are rapidly degraded
TCTGTGCTGG-39) and a 39 primer (59-TTTTTTTTTTT-
by the 26S proteasome, a multicatalytic protease complex (10,
TGT-39) and subcloned into pCRII (Invitrogen). This partial
11). Recent evidence shows that intracellular levels of cyclins
cDNA (298 bp) was shown by dideoxy DNA sequencing to
and cyclin dependent kinase inhibitors (12, 13), as well as other
contain the 59 and 39 primers. A cDNA library, from BayF3
growth regulatory proteins, such as p53 (14, 15), c-Jun (16),
cells growing in IL-3, was constructed in the phage vector
and IkBa (17), are regulated by ubiquitin-mediated proteol-
lZAP (Stratagene). Poly(A)1 mRNA used for library con-
ysis. It is also possible that ubiquitination alters a protein's
struction was prepared by the Fast Track mRNA Isolation Kit
function without affecting its metabolic stability (18).
(Invitrogen). The partial cDNA isolated by differential display
Little is known about the regulatory enzymes that determine
was labeled with [32P]dCTP by random prime labeling (27) and
which cellular proteins are specifically destroyed by ubiquitin-
used to screen 1 3 106 plaque-forming units from the library.
mediated proteolysis. Most evidence suggests that substrate
Three independent positive clones of different lengths that
specificity is determined by ubiquitin-conjugating enzymes
hybridized with the probe were isolated, and the corresponding
(19, 20). Recently, a large superfamily of genes encoding
plasmids were isolated from the phage clones. The longest
deubiquitinating enzymes was identified (21). Deubiquitinat-
cDNA clone was sequenced on both strands by the dideoxy
ing enzymes remove ubiquitin from intracellular protein con-
jugates by cleaving the amide linkage between the C terminus
DNA sequencing method (United States Biochemical).
of ubiquitin and either a-amino or «-amino groups of the
Abbreviations: IL-3, interleukin 3; IL-3R, IL-3 receptor; GST, gluta-
thione
S-transferase; ORF, open reading frame.
The publication costs of this article were defrayed in part by page charge
‡To whom reprint requests should be addressed at: Division of
payment. This article must therefore be hereby marked ‘‘
advertisement'' in
Pediatric Oncology, Dana–Farber Cancer Institute, 44 Binney Street,
accordance with 18 U.S.C. §1734 solely to indicate this fact.
Boston, MA 02115.
Cell Biology: Zhu
et al.
Proc. Natl. Acad. Sci. USA 93 (1996)
Northern Blot Analysis. RNA samples (10–30 mg) were
electrophoresed on denaturing formaldehyde gels and blotted
onto Duralon-UV membranes (Stratagene). The cDNA in-
serts, purified from agarose gels (Qiagen, Chatsworth, CA),
were radiolabeled (27) and hybridized for 1 hr to the filters in
a 68°C oven. Hybridized filters were finally washed at room
temperature in 0.13 SSC (13 SSC 5 0.15 M sodium chloridey
0.015 M sodium citrate, pH 7) and 0.1% SDS.
Deubiquitination Assay. The deubiquitination assay of ubiq-
uitin–b-galactosidase fusion proteins has been previously de-
scribed (21). A 1578-bp fragment from the wild-type
DUB-1
cDNA (corresponding to aa 1 to 526) and a cDNA containing
a missense mutation (C60S) were generated by polymerase
chain reaction (PCR) and inserted, in frame, into pGEX-2TK
(Pharmacia) downstream of the glutathione
S-transferase
(GST) coding element. Ub–Met–bgal was expressed from a
pACYC184-based plasmid. Plasmid-bearing
Escherichia coli
MC1061 cells were lysed and analyzed by immunoblotting with
anti-bgal antibodies (Cappel) and the enhanced chemilumi-
nescence system (Amersham).
Generation of Anti-DUB-1 Antiserum and Analysis of the
DUB-1 Polypeptide. A
DUB-1 antiserum was raised by inject-
ing a full-length GST–
DUB-1 fusion protein into a New
Zealand White rabbit and was affinity purified with a GST–
DUB-1 affinity matrix, as previously described (28).
In vitro
translation of the full length
DUB-1 polypeptide was per-
formed by standard procedures (Promega). Immunoblotting
was performed as previously described (29) using the affinity-
purified anti-
DUB-1 antiserum and enhanced chemilumines-
cence technology.
Heterologous Expression of DUB-1 in Bay
F3 Cells and Cell
Growth Analysis. The open reading frame (ORF) of
DUB-1
[or
DUB-1(C60S)] was generated by PCR using the following
39 (219 to 1) and 59-ATCTCGAGGTGTCCACAGGAGCCT-
GTGT-39 (1802 to 1781). The fragments (1637 bp) were
subcloned into the
Sma Iy
Xho I cloning sites of pMSG
(Pharmacia), which contains a mouse mammary tumor virus–
long terminal repeat inducible promoter and a
gpt selection
marker. Parental BayF3 cells were electroporated with vector
alone or with pMSG–
DUB-1 as previously described (25).
After 3 days in IL-3 medium, the cells were selected in IL-3
medium containing 250 mgyml xanthine, 15 mgyml hypoxan-
thine, 10 mgyml thymidine, 2 mgyml aminopterin, and 25
mgyml mycophenolic acid.
Gpt-resistant subclones were iso-
lated by limiting dilution.
DUB-1 expression was induced by
adding 0.1 mM dexamethasone (diluted from 10 mM stock in
ethanol). Cell proliferation and cell viability were measured by
trypan blue exclusion (25).
Analysis of Cell Cycle. Cell cycle analysis was performed by
fluorescence-activated cell sorter, as previously described (30).
The percentage of cells in each phase of the cell cycle was
determined by analyzing data with the computer programCELLFIT (Becton Dickinson).
DUB-1 Is a Hematopoietic-Specific Immediate Early Gene
Encoding a Deubiquitinating Enzyme. BayF3 is a murine
FIG. 1. Sequence and homologies of the
DUB-1 cDNA. (
A) Nucle-
lymphocyte cell line that depends on IL-3 for growth and
otide and predicted amino acid sequence of
DUB-1. Underlined se-
viability (24, 30, 31). By comparing mRNA from IL-3-deprived
quences are copies of a conserved motif shown by Shaw and Kamen (32)
and IL-3-stimulated BayF3 cells (22, 23), we initially isolated
to confer message instability and which are found in the 39 untranslated
an IL-3 inducible, immediate early cDNA fragment (
DUB-1).
regions of many mitogen-induced, immediate early mRNAs. A consensus
The full-length 2674-bp
DUB-1 cDNA was subsequently iso-
polyadenylylation signal is double underlined. The sequence of the
lated and found to contain a 1581-bp ORF (Fig. 1
A). There are
murine
DUB-1 cDNA has been assigned GenBank no. 24133 U41636. (
B)
two stop codons within the 183 bp of 59 untranslated region.
Sequence homologies between yeast Doa4 (21), human
Tre-2 (33),
murine Unp (34), and murine
DUB-1. Alignment of
DUB-1 with human
In addition, we isolated a murine genomic clone that contains
c-
myc is also shown. The homologous domain of c-
myc contains the
a TATA box at position 2321 and an IL-3 inducible enhancer
nuclear localization sequence PAAKRAKLD (35) but not the c-
myc
(Y.Z., unpublished data).
DNA binding domain.
Cell Biology: Zhu et al.
Proc. Natl. Acad. Sci. USA 93 (1996)
The DUB-1 ORF is predicted to encode a polypeptide of 526
aa (59 kDa). Comparison of the DUB-1 protein sequence with
entries in GenBank data base (3y96) detected significant
similarity with several deubiquitinating enzymes, including
Tre-2 (33, 36), Unp (34), and Doa4 (21). The sequence
similarity was largely restricted to the conserved Cys and His
boxes previously identified for this enzyme superfamily (Fig.
1B) (21). These elements probably help form the enzyme
active site (21). The likely active site nucleophile is a cysteine
residue in the Cys box that is found in all known family
members (21) and is also present in DUB-1 (Cys60). The 39
untranslated region of the DUB-1 cDNA contained two AT-
TTA sequences, located in A 1 T rich domains. The AUUUA
sequence, found in the 39 untranslated regions of many im-
mediate early mRNAs, may play a role in DUB-1 mRNA
turnover (32). The DUB-1 mRNA was detected in multiple
hematopoietic cell lines, but not in nonhematopoietic cell lines
or tissues from adult mice (data not shown).
DUB-1 Encodes a Functional Deubiquitinating Enzyme. In
order to determine whether DUB-1 has deubiquitinating ac-
tivity, we expressed DUB-1 as a GST fusion protein. The
DUB-1 ORF was subcloned into the bacteria expression vector,
pGEX. pGEX–DUB-1 was co-transformed into E. coli
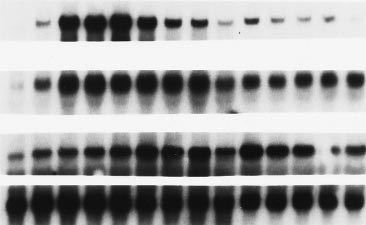
FIG. 3. DUB-1 mRNA levels are induced by IL-3 in early G1 phase,
(MC1061) with a plasmid expressing the protein Ub–Met–b
followed by a rapid decline. BayF3-EPO-R cells were arrested in early
gal, in which ubiquitin is fused to the N terminus of b-galac-
G1 phase by growth factor starvation for 12 hr and were restimulated
tosidase. As shown by immunoblot analysis (Fig. 2), two
with IL-3 to enter the cell cycle synchronously. Total RNA (10 mg per
independent cDNA clones encoding GST–DUB-1 fusion pro-
lane) extracted from cells at the indicated time (in hours) was
tein resulted in cleavage of Ub–Met–b gal (lanes 3, 4, and 7)
subjected to Northern blot analysis with the indicated cDNA probes.
The different cell cycle phases were determined by flow-cytometric
comparable to that observed with Ubp1, a known yeast
analysis of cellular DNA content.
deubiquitinating enzyme (21) (lane 1). As controls, cells with
the pGEX vector (lane 5) or pBluescript vector with a non-
Continuous Expression of DUB-1 Arrests Cellular Growth.
transcribed DUB-1 insert (lane 2) failed to cleave Ub–Met–b
Our initial attempts to obtain stable cell lines that consti-
gal. A mutant DUB-1 polypeptide, containing a C60S muta-
tutively express DUB-1 were unsuccessful. Because DUB-1
tion, was unable to cleave the Ub–Met–b gal substrate (lane 6).
expression is normally turned off after only a brief period of
Expression of GST-DUB-1 in bacterial cells containing the
synthesis (Fig. 3), we reasoned that continuous expression of
Ub–Leu–b gal substrate showed greatly reduced levels of b gal
DUB-1 mRNA might somehow interfere with cell growth
activity (data not shown). The Leu–b gal product, unlike
andyor viability. We therefore expressed DUB-1 in BayF3
Met–b gal or the respective Ub–b gal fusions, is short lived in
cells using an inducible promoter (Fig. 4). Twelve gpt-
E. coli (37). This result strongly suggests that DUB-1 cleaves
resistant BayF3 subclones were generated after transfection
Ub–Leu–b gal specifically at the C terminus of the ubiquitin
with either pMSGyDUB-1 or mutant pMSGyDUB-1(C60S),
moiety. Taken together, these results demonstrate that DUB-1
which encodes the inactive enzyme. Dexamethasone (0.1
has deubiquitinating activity and that Cys 60 is critical for its
mM) induced DUB-1 mRNA in all transfected cells, but not
thiol protease activity.
in parental or mock-transfected cells (data not shown).
DUB-1 mRNA Levels Are Induced by IL-3 in Early G1 Phase,
Dexamethasone induced expression of the DUB-1 protein
Followed by a Rapid Decline. BayF3 cells arrest in early G1
(Fig. 4A, lane 2) or DUB-1(C60S) protein (lane 4) in trans-
phase when deprived of IL-3 for 12 hr and can be induced to
fected BayF3 cells to levels comparable with those observed
reenter the cell cycle synchronously by readdition of growth
during IL-3-induced expression from the endogenous DUB-1
factor (30). The 3.1-kb DUB-1 mRNA appeared 30 to 60 min
gene (data not shown). These proteins had the same electro-
after addition of IL-3 (Fig. 3) but rapidly decreased in abun-
phoretic mobility (59 kDa) as full-length DUB-1 polypeptide
dance before the completion of G1 phase. DUB-1 mRNA levels
synthesized by in vitro translation (lane 5). After dexametha-
were superinduced with IL-3 plus cycloheximide (data not
sone induction, cells expressing DUB-1 failed to proliferate in
shown), defining DUB-1 as an immediate early gene. Induction
IL-3 medium (Fig. 4B). In contrast, dexamethasone-induced
of DUB-1 mRNA was similar to that of c-myc, although c-myc
cells expressing DUB-1 (C60S) proliferated normally in IL-3.
mRNA levels remained elevated throughout G1 phase. Cyclin
Importantly, while dexamethasone induction of wild-type
D2 mRNA accumulated later in G1 phase as previously
DUB-1 inhibited cellular proliferation, as measured by total
described (38).
cell number, it had little effect on cellular viability (Table 1).
The BayF3 subclones that were induced with dexamethasone to
express either wild-type DUB-1 or DUB-1(C60S) remained viable
in IL-3. Cells underwent apoptosis only after removal of IL-3.
To test the possibility of nonspecific toxicity caused by pro-
longed expression of active DUB-1 enzyme, we stopped DUB-1
synthesis in cells transfected with the wild-type DUB-1 construct
by removal of dexamethasone at day 7. Cells resumed normal
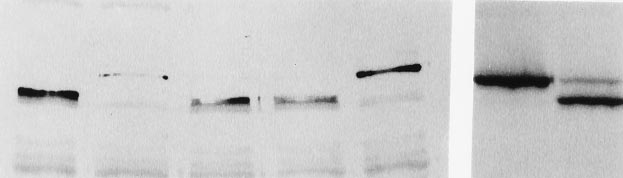
FIG. 2. DUB-1 encodes a functional deubiquitinating enzyme.
proliferation within 48 hr following dexamethasone with-
Deubiquitination of ubiquitin-b-galactosidase (Ub–Met–bgal) fusion
drawal (Fig. 4C). To provide further evidence against non-
proteins expressed in bacteria. Shown is a Western blot using anti-
specific toxicity, we induced DUB-1 expression in murine
bgal antiserum. Co-expressed plasmids were pGEX–Ubp1 (lane 1)
3T3 fibroblasts (data not shown). Normal cell proliferation
(21), pBluescriptyDUB-1 (DUB-1 is not expressed) (lane 2), pGEX–
DUB-1.1 (lanes 3 and 7), pGEX–DUB-1.2 (lane 4), pGEX(vector)
was observed for these cells, indicating that growth suppres-
(lane 5), and pGEX–DUB-1(C68S) (lane 6).
sion by DUB-1 is cell-type specific.
Cell Biology: Zhu et al.
Proc. Natl. Acad. Sci. USA 93 (1996)
Table 1. Percent viable BayF3 cells
DUB-1 (WT)
DUB-1 (C60S)
WT, wild type; DEX, dexamethasone.
Normally, DUB-1 mRNA levels rise soon after IL-3 addition
during the early G1 phase of the cell cycle, followed by a rapid
decline. When DUB-1 mRNA levels are maintained by con-
tinuous synthesis from a dexamethasone-inducible promoter,
BayF3 cells arrest in the G1 phase of the cell cycle. These data
indicate that DUB-1 expression is tightly regulated and that
DUB-1 may play a role in cytokine-induced cell proliferation.
Deubiquitinating enzymes studied in yeast have multiple
functions (21). Some deubiquitinating enzymes, such as Ubp2,
can apparently remove ubiquitin from ubiquitin-conjugated
substrates prior to proteasome-substrate binding, thereby
slowing the turnover of such proteins (39). Other deubiquiti-
nating enzymes, such as Doa4, may remove ubiquitin from
proteasome-bound degradation products, allowing recycling
of ubiquitin and proteasomes and thereby promoting further
protein degradation (21). Ubiquitin must also be cleaved from
precursor forms by deubiquitinating enzymes. Finally, dynamic
ubiquitination events may serve as reversible regulatory
switches (40, 41).
Failure to turn off expression of DUB-1 presumably, as in
our experiments, may cause G1 arrest by preventing the
degradation of growth-inhibitory proteins, such as cyclin de-
pendent kinase inhibitors, or by promoting the degradation of
growth-permissive proteins, such as G1 cyclins. Alternatively,
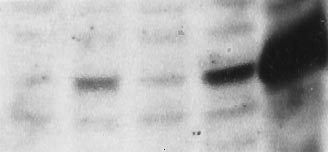
FIG. 4. Continuous expression of DUB-1 results in growth sup-
pression. (A) Immunoblot analysis of steroid-induced DUB-1
polypeptide. Lysates (100 mg of total protein) from the indicated
cells were electrophoresed in 10% SDS polyacrylamide gels and
blotted with affinity-purified anti-DUB-1 antibody (1:1000). (B)
BayF3-DUB-1 (open symbols) or BayF3-DUB-1(C60S) (filled sym-
bols) were cultured in IL-3 medium with (‚, Œ) or without (h, ■)
dexamethasone (0.1 mM). Cell number was calculated by the trypan
blue exclusion technique. (C) BayF3 cells, transfected with wild-type
DUB-1, were grown in IL-3 with (‚) or without (h) dexamethasone
for 6 days. Dexamethasone-treated cells were washed, replated in
IL-3 medium (without dexamethasone) on day 7 (arrow), and
cultured for an additional 6 days (‚).
Cell cycle analysis demonstrated that the majority of
BayF3 cells were arrested in the G1 phase of the cell cycle
following dexamethasone induction of DUB-1 (Fig. 5). This
concentration of dexamethasone (0.1 mM) slightly reduced
IL-3 dependent proliferation of parental BayF3 cells or
DUB-1(C60S)-expressing cells (to 80% of maximum), but it
FIG. 5. Forced expression of DUB-1 results in growth arrest in the
completely blocked proliferation of the wild-type DUB-1-
G1 phase of the cell cycle. The indicated cell lines were grown for 48
expressing cells.
hr (5 3 105 cellsyml) with or without dexamethasone (0.1 mM). The
cells were stained with propidium iodide and analyzed by flow
cytometry. The percentage of cells in G1, S, and G2yM were: DUB-1
2 dexamethasone (32%, 61%, 7%), 1 dexamethasone (82%, 14%,
4%); DUB-1(C60S) 2 dexamethasone (35%, 57%, 8%), 1 dexameth-
In the present work, we describe a novel murine immediate
asone (35%, 57%, 8%). Data shown are representative of at least three
early gene that encodes a deubiquitinating enzyme, DUB-1.
separate dexamethasone induction experiments.
Cell Biology: Zhu et al.
Proc. Natl. Acad. Sci. USA 93 (1996)
DUB-1 may specifically regulate proteolysis (or the ubiquiti-
10. Peters, J.-M., Cejka, Z., Harris, J. R., Kleinschmidt, J. A. &
nation state) of a protein in an IL-3-specific signal transduction
Baumeister, W. (1993) J. Mol. Biol. 234, 932–937.
pathway. Identification of the specific substrates of DUB-1
11. Peters, J.-M. (1994) Trends Biochem. Sci. 19, 377–382.
should help elucidate its mechanism of growth suppression.
12. Glotzer, M., Murray, A. W. & Kirschner, M. W. (1991) Nature
Constitutive expression of wild-type DUB-1 does not suppress
(London) 349, 132–138.
13. Pagano, M., Tam, S. W., Theodoras, A. M., Beer-Romero, P.,
the growth of murine 3T3 fibroblasts (data not shown). This
Del Sal, G., Chau, V., Yew, P. R., Draetta, G. F. & Rolfe, M.
suggests that the growth suppression by DUB-1 might be
(1995) Science 269, 682–686.
specific to hematopoietic cells, the only cell types in which
14. Scheffner, M., Huibregtse, J. M., Vierstra, R. D. & Howley, P. M.
DUB-1 is normally expressed. This may reflect the existence
(1993) Cell 75, 495–505.
of a hematopoietic-specific substrate(s) of DUB-1, a hemato-
15. Chowdary, D. R., Dermody, J. J., Jha, K. K. & Ozer, H. L. (1994)
poietic cell-restricted DUB-1 cofactor, or a higher threshold of
Mol. Cell. Biol. 14, 1997–2003.
resistance to continuous DUB-1 expression in other cell types.
16. Treier, M., Staszewski, L. M. & Bohmann, D. (1994) Cell 78,
Interestingly, we have isolated three additional genes whose
17. Chen, Z., Hagler, J., Palombella, V. J., Melandri, F., Scherer, D.,
predicted products show high sequence similarity to DUB-1
Ballard, D. & Maniatis, T. (1995) Genes Dev. 9, 1586–1597.
(approximately 80% amino acid identity) (Y.Z., unpublished
18. Hochstrasser, M. (1995) Curr. Opin. Cell Biol. 7, 215–223.
observations). These genes are presumably DUB subfamily
19. King, R. W., Peters, J.-M., Tugendreich, S., Rolfe, M., Hieter, P.
members, and we refer to them as DUB-2 through DUB-4.
& Kirschner, M. W. (1995) Cell 81, 279–288.
They may be induced by different growth factors andyor may
20. Tugendreich, S., Tomkiel, J., Earnshaw, W. & Hieter, P. (1995)
deubiquitinate different intracellular substrates.
Cell 81, 261–268.
We hypothesize that, like other immediate early gene prod-
21. Papa, F. R. & Hochstrasser, M. (1993) Nature (London) 366,
ucts, DUB-1 plays a role in integrating extracellular signals with
22. Liang, P. & Pardee, A. B. (1992) Science 257, 967–971.
cellular growth and cell cycle progression. Our data suggest
23. Liang, P., Averboukh, L. & Pardee, A. B. (1993) Nucleic Acids
that turning off DUB-1 expression after a rapid burst of
Res. 21, 3269–3275.
IL-3-induced synthesis is crucial for hematopoietic cell prolif-
24. Palacios, R. & Steinmetz, M. (1985) Cell 41, 727–734.
eration. It is possible that after cytokine induction, cells only
25. D'Andrea, A. D., Yoshimura, A., Youssoufian, H., Zon, L. I.,
turn off DUB-1 under specific conditions, e.g., adequate
Koo, J.-W. & Lodish, H. F. (1991) Mol. Cell. Biol. 11, 1980–1981.
nutrient availability, thereby providing a mechanism by which
26. Chirgwin, J. M., Przybyla, A. E., MacDonald, R. G. & Rutter,
a cell could halt cell cycle progression following exposure to a
W. J. (1979) Biochemistry 18, 5294–5299.
mitogenic signal. Several other examples of mitogen-induced
27. Feinberg, A. P. & Vogelstein, B. (1983) Anal. Biochem. 132,
negative regulators have recently been demonstrated. MKP-1
28. Yamashita, T., Barber, D., Zhu, Y., Wu, N. & D'Andrea, A. D.
(42) and PAC1 (43) are mitogen-induced threonineytyrosine
(1994) Proc. Natl. Acad. Sci. USA 91, 6712–6716.
phosphatases that inactivate mitogen activating protein kinase.
29. Barber, D. L. & D'Andrea, A. D. (1994) Mol. Cell. Biol. 14,
p21 is a mitogen-induced inhibitor of cyclinycdk complexes
(44). DUB-1 is the first enzyme of the ubiquitin system directly
30. Carroll, M., Zhu, Y. & D'Andrea, A. D. (1995) Proc. Natl. Acad.
implicated in cytokine-regulated growth control.
Sci. USA 92, 2869–2873.
31. Liboi, E., Carroll, M., D'Andrea, A. D. & Mathey-Prevot, B.
(1993) Proc. Natl. Acad. Sci. USA
We thank Peng Liang, Bernard Mathey-Prevot, David Pellman,
32. Shaw, G. & Kamen, R. (1986) Cell 46, 649–667.
Mark Ewen, Joan Ruderman, Chuck Stiles, and members of the
33. Nakamura, T., Hillova, J., Mariage-Samson, R., Onno, M.,
D'Andrea laboratory for helpful discussions. We thank Barbara Keane
Huebner, K., Cannizzaro, L. A., Boghosian-Sell, L., Croce, C. M.
for preparation of the manuscript. This work was supported by
& Hill, M. (1992) Oncogene 7, 733–741.
National Institutes of Health Grant RO1 DK 43889-01 (to A.D.D.).
34. Gupta, K., Copeland, N. G., Gilbert, D. J., Jenkins, N. A. & Gray,
and Grant GM46904 (to M.H.). Y.Z. is a Fellow of the Leukemia
D. A. (1993) Oncogene 8, 2307–2310.
Society of America. A.D.D. is a Lucille P. Markey Scholar and a
35. Dang, C. V. & Lee, W. M. F. (1988) Mol. Cell. Biol. 8, 4048–4054.
Scholar of the Leukemia Society of America, and this work was also
36. Onno, M., Nakamura, T., Mariage-Samson, R., Hillova, J. & Hill,
supported in part by a grant from the Lucille P. Markey Charitable
M. (1993) DNA Cell. Biol. 12, 107–118.
37. Tobias, J. W., Shrader, T. E., Rocap, G. & Varshavsky, A. (1991)
Science 254, 1374–1377.
1. Ihle, J. N. (1993) in Interleukins: Molecular Biology and Immu-
38. Matsushime, H., Roussel, M. F., Ashmun, R. A. & Sherr, C. J.
nology, ed. Kishimoto, T. (Karger, Basel), pp. 65–106.
(1991) Cell 65, 701–713.
2. Kitamura, T., Sato, N., Arai, K.-i. & Miyajima, A. (1991) Cell 66,
39. Baker, R. T., Tobias, J. W. & Varshavsky, A. (1992) J. Biol. Chem.
3. Sato, N., Sakamaki, K., Terada, N., Arai, K. & Miyajima, A.
40. Paolini, R. & Kinet, J. P. (1993) EMBO J. 12, 779–786.
(1993) EMBO J. 12, 4181–4189.
41. Corsi, D., Galluzzi, L., Crinelli, R. & Magnani, M. (1995) J. Biol.
4. Murray, A. (1995) Cell 81, 149–152.
Chem. 270, 8928–8935.
5. Sherr, C. J. & Roberts, J. M. (1995) Genes Dev. 9, 1149–1163.
42. Sun, H., Charles, C. H., Lau, L. F. & Tonks, N. K. (1993) Cell 75,
6. Hershko, A. (1988) J. Biol. Chem. 263, 15237–15240.
7. Hershko, A. & Ciechanover, A. (1992) Annu. Rev. Biochem. 61,
43. Ward, Y., Gupta, S., Jensen, P., Wartmann, M., Davis, R. J. &
Kelly, K. (1994) Nature (London) 367, 651–654.
8. Finley, D. & Chau, V. (1991) Annu. Rev. Cell Biol. 7, 25–69.
44. Michieli, P., Chedid, M. M., Lin, D., Pierce, J. H., Mercer, W. E.
9. Ciechanover, A. (1994) Cell 79, 13–21.
& Givol, D. (1994) Cancer Res. 54, 3391–3395.
Source: http://goby.compbio.cs.cmu.edu/NC/Survey/USP/Zhu96.pdf
Payments and Quality of Ante-NatalCare in Two Rural Districts ofTanzania Paper 4 from the Ethics, Payments, and P.O. Box 33223, Dar es Salaam, Tanzania Maternal Survival Project 157 Mgombani Street, Regent EstateTel: +255 (0) 22 2700083 / 2772556Fax: +255 (0) 22 2775738 Paula Tibandebage, Maureen Mackintosh, Tausi Kida,
Drug Therapies There are many prescription drugs available to treat MS. There are disease modifying drugs that affect the long-term course of MS and other medications to treat a particular symptom. Disease Modifying Treatments (DMTs) There are number of disease-modifying treatments (DMTs) which can reduce the frequency and severity of clinical relapses. The National Institution for Care Excellence (NICE) published guidelines in October 2014 for the management of multiple sclerosis. In these guidelines a relapse can be diagnosed by a GP or neurologist if: The person with MS has developed new symptoms or has a worsening of existing symptoms, and these symptoms have lasted for more than 24 hours in the absence of infection of any other cause after a stable period of a least 1 month. The following DMTs are available for treatment of relapsing remitting multiple sclerosis, CIS andwho are still experiencing relapses, and where these relapses are the predominant cause of their increasing disability :- Avonex - a weekly injection into the muscle.