Kamagra gibt es auch als Kautabletten, die sich schneller auflösen als normale Pillen. Manche Patienten empfinden das als angenehmer. Wer sich informieren will, findet Hinweise unter kamagra kautabletten.
Skeletalmusclejournal.com
Knoblauch et al. Skeletal Muscle 2013, 3:22http://www.skeletalmusclejournal.com/content/3/1/22
Mice with RyR1 mutation (Y524S) undergohypermetabolic response to simvastatin
Mark Knoblauch†, Adan Dagnino-Acosta† and Susan L Hamilton*
Background: Statins are widely used drugs for the treatment of hyperlipidemia. Though relatively safe, someindividuals taking statins experience rhabdymyolysis, muscle pain, and cramping, a condition termed statin-inducedmyopathy (SIM). To determine if mutations in the skeletal muscle calcium (Ca2+) release channel, ryanodinereceptor type 1 (RyR1), enhance the sensitivity to SIM we tested the effects of simvastatin, the statin that producesthe highest incidence of SIM in humans, in mice with a mutation (Y524S, ‘YS') in RyR1. This mutation is associatedwith malignant hyperthermia in humans. Exposure of mice with the YS mutation to mild elevations inenvironmental temperature produces a life-threatening hypermetabolic response (HMR) that is characterized byincreased oxygen consumption (VO2), sustained muscle contractures, rhabdymyolysis, and elevated core bodytemperature.
Methods: We assessed the ability of simvastatin to induce a hypermetabolic response in the YS mice using indirectcalorimetry and to alter Ca2+ release via RyR1 in isolated flexor digitorum brevis (FDB) fibers from WT and YS miceusing fluorescent Ca2+ indicators. We also tested the ability of 5-aminoimidazole-4-carboxamide ribonucleoside(AICAR) to protect against the simvastatin effects.
Results: An acute dose of simvastatin triggers a hypermetabolic response in YS mice. In isolated YS muscle fibers,simvastatin triggers an increase in cytosolic Ca2+ levels by increasing Ca2+ leak from the sarcoplasmic reticulum (SR).
With higher simvastatin doses, a similar cytosolic Ca2+ increase occurs in wild type (WT) muscle fibers. Pre-treatmentof YS and WT mice with AICAR prevents the response to simvastatin.
Conclusions: A mutation in RyR1 associated with malignant hyperthermia increases susceptibility to an adverseresponse to simvastatin due to enhanced Ca2+ release from the sarcoplasmic reticulum, suggesting that RyR1mutations may underlie enhanced susceptibility to statin-induced myopathies. Our data suggest that AICAR may beuseful for treating statin myopathies.
Keywords: Statin-induced myopathy, Simvastatin, RyR1, Myopathy, Calcium signaling
results in a condition termed statin-induced myopathy
Statins (3-hydroxy-3-methylglutaryl coenzyme-A (HMG-
(SIM) A mechanism to explain the underlying cause
CoA) reductase inhibitors) are cholesterol-lowering drugs
of SIM has yet to be elucidated.
that have proven effective in decreasing low-density lipo-
One emerging theory of SIM has centered on statins'
protein (LDL) levels and improving overall health For
potential to modulate intramyofiber calcium (Ca2+)
the majority of patients, statins are well tolerated with few
homeostasis [. This theory stems in part from the
side effects. However, up to 10% of patients on a statin regi-
finding that the direct application of simvastatin to healthy
men display muscle-related symptoms including soreness,
human myofibers triggers a significant increase in cyto-
fatigue, and an increase in circulating levels of muscle-
solic Ca2+ . The sudden release of Ca2+ in response to
specific proteins (for example creatine kinase (CK)) that
direct application of statins in vitro has been suggested tooriginate from both mitochondria and the sarcoplasmic
* Correspondence:
reticulum (SR) , the predominant Ca2+ storage or-
†Equal contributors
ganelle within the myofiber. The potential involvement of
Department of Molecular Biology and Biophysics, Baylor College of Medicine,1 Baylor Plaza, Houston, TX 77030, USA
2013 Knoblauch et al.; licensee BioMed Central Ltd. This is an Open Access article distributed under the terms of theCreative Commons Attribution License which permits unrestricted use,distribution, and reproduction in any medium, provided the original work is properly cited.
Knoblauch et al. Skeletal Muscle 2013, 3:22
the SR in statin-induced Ca2+ release is particularly
studies involving injection into mice for indirect calor-
intriguing given the recent findings that mutations in
imetry, simvastatin powder was dissolved in dimethyl sulf-
ryanodine receptor type 1 (RyR1), the Ca2+ release
oxide (DMSO). For single-fiber perfusion work, a 12 mM
channel of the SR, may underlie some instances of SIM
simvastatin stock was prepared in 10% EtOH similar to
[Mutations in RyR1 are known to produce malig-
previous studies []. After adjusting the pH to 7.0, the
nant hyperthermia (MH), a life-threatening condition
solution was brought up to 12 mM concentration in
where uncontrolled release of Ca2+ within the myofiber
Tyrode's solution containing 121 mM NaCl, 5 mM KCl,
is triggered by exposure to certain volatile inhalants,
1.8 mM CaCl2, 500 μM MgCl2, 400 μM NaH2PO4, 100
elevated temperature, or exercise ]. This uncon-
μM EDTA, 5.5 mM glucose, and 24 mM NaHCO3. Separ-
trolled release of Ca2+ results in sustained muscle contrac-
ately, a vehicle-only stock was prepared identically but
tions, elevated core temperature, rhabdomyolysis and, if
without the addition of simvastatin. These prepared stocks
unabated, death [].
were aliquoted and frozen at −80°C until use.
At present our understanding of the link between
RyR1 mutations and statin myopathies has been limited
Indirect calorimetry monitoring of VO2 max
to in vitro work with muscle biopsies. Metterlein et al.
Those YS and WT mice used to determine the effects
found that biopsied muscle from MH-sensitive swine ex-
of statin dosing in vivo were removed from their cage,
hibit contraction upon exposure to statins in vitro
weighed, and injected IP with an 30-80 mg/kg dose of ei-
Similarly, Guis et al. found that muscle biopsies from
ther simvastatin dissolved in DMSO or DMSO alone (‘ve-
seven of nine human subjects exhibiting the signs of SIM
hicle'). The mice were then returned to their cages for
expressed abnormal in vitro contracture tests (IVCT) used
30 minutes, after which they were placed individually
to screen for susceptibility to MH
into an environmental chamber at 32°C containing indir-
These in vitro findings combined with evidence that
ect calorimetry chambers (Oxymax System, Columbus
simvastatin modifies Ca2+ homeostasis suggest that
Instruments, Columbus, OH, USA), which allowed for
RyR1 mutations may underlie enhanced susceptibility to
monitoring of maximum oxygen consumption (VO2
SIM. We developed a mouse model (Y524S, ‘YS') with a
max (mL/kg/min)). Separately, to evaluate the effect-
RyR1 knock-in mutation of tyrosine 524 to serine
iveness of a pharmaceutical agent shown previously to
which in humans (Y522S) is associated with MH
prevent heat-induced HMR response in YS mice [
Mice homozygous for the mutation die at birth, while
additional YS mice were injected IP with a 600 mg/kg
heterozygous YS mice exhibit a hypermetabolic response
dose of 5-aminoimadazole-4-carboxamide ribonucleo-
(HMR) to elevated (37°C) temperature, volatile anes-
side (AICAR) 20 minutes after simvastatin injection.
thetics, or exercise in a warm environment. These miceare a valuable tool for studying some RyR1-associated
disorders. The purpose of the present study was to de-
For mice destined for single-fiber Ca2+ study, the flexor
termine whether mice with this RyR1 mutation (Y524S)
digitorum brevis (FDB) muscle was removed and imme-
display HMR when given simvastatin and to evaluate the
diately placed into Dulbecco's modified Eagle's medium
effects of simvastatin on intramyofiber Ca2+ homeostasis.
(DMEM) containing 3 mg/mL collagenase and 10% (v/v)fetal bovine serum. After a 2-hour incubation at 37°C,
whole FDB muscles were transferred to 1 mL of DMEM
Animal care and handling
and plunged ten times through a 1 mL pipette tip to
All procedures were approved by the Institutional
separate individual fibers. Next, 150 μL of DMEM
Animal Care and Use Committee at Baylor College of
containing separated FDB fibers was placed onto a 25 mm
Medicine, Houston, TX, USA. As previously described,
glass coverslip that had been incubated for 2 hours with
male RyR1Y524S/WT (‘YS') mice were developed and used
20 μg/mg of laminin in PBS and then subjected to
in conjunction with wild type (WT) littermate controls at
two washes in PBS and a final wash in DMEM. Prior
8 to 10 weeks of age. Mice were maintained on a 12:12
to use, plated fibers were incubated overnight at 37°C in
light:dark cycle, had ad libitum access to water and stand-
DMEM containing antibiotic-antimycotic (Gibco, Carlsbad,
ard mouse chow, and were limited to normal cage activity
only. All mice were sacrificed at the same time of day,consisting of cervical dislocation after anesthetization
Isolated fiber preparation and imaging
under isoflurane.
To assess the sensitivity to simvastatin, after the over-night incubation the fibers were next incubated for 1
Statin preparation
hour at room temperature in either DMEM containing
Simvastatin was purchased from the manufacturer (LKT
(10 μM) Fura-2 acetoxymethyl ester (Fura-2 AM) or 30
Laboratories, St Paul, MN, USA) in powder form. For
minutes in DMEM containing (5 μM) Mag-fluo-4, with
Knoblauch et al. Skeletal Muscle 2013, 3:22
(20 μM) contraction-inhibitor 4-methyl-N-(phenylmethyl)
the mice to be euthanized prior to a full body contrac-
benzenesulfonamide (BTS). Fibers were placed in a
tion and death. To determine if statins also trigger an
temperature controlled chamber (Dagan Corporation,
HMR response, we injected mice with an acute dose of
Minneapolis, MN, USA) on the stage of an inverted epi-
simvastatin (IP 30 to 80 mg/kg) and placed the mice in
fluorescence microscope (Nikon Inc, Melville, NY, USA)
the chamber (32°C, a thermoneutral temperature that
and warmed to 32°C over a 5-minute period in Tyrode's
does not trigger HMR in the untreated YS mice) of the
solution. Fluorescence emission was captured using a
indirect calorimeter and measured VO2 as a function
high speed, digital QE CCD camera (TILL Photonics,
of time after injection. All YS mice injected with 60
Pleasanton, CA, USA). Each fiber was tested against a sin-
or 80 mg/kg simvastatin exhibited subsequent signs of
gle dose of simvastatin, and peak fluorescence values were
HMR, which included increased VO2 (Figure A), severe
averaged across all fibers per group for each concentration.
muscle contractures and increased heat production. Afterinjection with simvastatin, a significantly higher peak VO2
Simvastatin sensitivity and AICAR effectiveness in
occurred in YS mice receiving 60 mg/kg (P <0.05) and
80 mg/kg (P <0.001) doses when compared against
To determine the effects of simvastatin dosing, YS and
YS mice injected with the vehicle. Figure shows
WT fibers loaded with Fura-2 AM were perfused for
the dose–response curve for peak VO2 as a function
2 minutes in warmed (32°C) Tyrode's solution for record-
of simvastatin dose in the YS mice.
ing of baseline Ca2+ levels, followed by a 3-minute expos-
To determine if the statin-induced HMR event was
ure at specified doses of simvastatin. Separately, isolated
similar to heat-induced HMR in the YS mice, we
fibers used to test AICAR's effectiveness at preventing the
injected the YS mice with 80 mg/kg of simvastatin
statin-modulated change in Ca2+ were pre-incubated in
followed by 600 mg/kg of AICAR, which we have previ-
1 mM AICAR in conjunction with the 1-hour incuba-
ously shown to prevent temperature-induced HMR in
tion in DMEM/Fura-2 AM before exposure to 500 μM
the YS mice by decreasing Ca2+ leak from RyR1
and 1 mM simvastatin in the YS and WT, respectively.
AICAR eliminated the statin-associated HMR in YS
Fura-2 fluorescence was recorded and converted to cyto-
mice by preventing the significant (P <0.01) increase in
solic Ca2+ values as previously reported
VO2 that occurs in YS mice not receiving the AICARtreatment (Figure ).
4-CMC-induced Ca2+ store depletion in isolated fibersTo evaluate the effects of simvastatin on SR Ca2+ storedepletion, isolated fibers were exposed to 4-chloro-m-
Myofibrillar Ca2+ leak is more sensitive to simvastatin in
cresol (4-CmC) immediately after 3 minutes of incuba-
YS compared to WT muscle fibers
tion in 500 μM simvastatin. 4-CmC was applied to either
The strong protective effect of AICAR on the simva-
YS or WT fibers at the dose found to induce maximal
statin response of the YS mice suggests that statin-
Ca2+ release without causing death of the individual
induced HMR in these mice is likely due to altered Ca2+
fibers, which we determined to be 1 mM in the YS and
handling within the myofiber. We tested the effects of
2.5 mM in the WT mice.
simvastatin in isolated FDB fibers of YS and WT miceusing the fluorescent dye Fura-2 to assess changes in
Statistical analysis
cytosolic Ca2+ concentrations. We found that simva-
A Student's t-test was used for comparison between
statin triggered higher cytosolic Ca2+ levels in YS fibers
groups to test significance values of P <0.05 (*), P <0.01
at lower concentrations (500 μM (P <0.001) and 750 μM
(**), and P <0.001 (***). Dose–response curves were fit
(P <0.01)) than in WT FDB fibers (Figure As previ-
using 4-parameter (oxygen consumption (VO2)) or 3-
ously shown with human fibers WT fibers displayed
parameter (single-fiber dose–response) Hill function
increased Ca2+ in response to higher doses of simva-
curves in SigmaPlot, version 12.0 (Systat Software,
statin (1.5 mM (P <0.01)). The concentration response
San Jose, CA, USA). YS data was additionally fitted
curves in the YS and WT mice were best fit using a Hill
with a biphasic function using GraphPad Prism, ver-
function (3-parameter) with a resulting EC50 of 0.6 mM
sion 6 (GraphPad Software, La Jolla, CA, USA).
in the YS and 0.9 mM in the WT mice. Since the YS fi-bers are from heterozygous mice, the Ca2+ response re-
flects the heterogeneous response from a mixture of
Simvastatin triggers HMR in YS mice
mutant channels (in various combinations of mutation
We previously demonstrated that changes in VO2 could
and WT subunits) and WT channels. Using a 2-site
be used to detect the HMR response in the YS mice ex-
model, we obtain EC50s of 0.4 and 0.9 mM. Ca2+ con-
posed to elevated environmental temperatures This
centrations were calculated from the Fura-2 fluorescence
approach allows early detection of the HMR and allows

Knoblauch et al. Skeletal Muscle 2013, 3:22
Figure 1 An acute dose of simvastatin at 32°C results in higher peak VO2 levels in YS compared to WT mice. (A) IP injection ofsimvastatin triggers significantly higher peak VO2 values at 60 mg/kg (P <0.05) and 80 mg/kg (P <0.001) compared to vehicle-only injection.
(B) Curve-fit of increasing simvastatin doses in YS mice. (C) Pre-treatment with 600 mg/kg AICAR results in significantly (P <0.01) lower peak VO2values when administered 20 minutes after simvastatin treatment in YS mice. (D) Representative VO2 tracings of YS mice receiving 80 mg/kg ofsimvastatin show increasingly higher VO2 values than YS mice treated with both 80 mg/kg simvastatin and 600 mg/kg AICAR, vehicle-only(DMSO), or WT mice treated with 80 mg/kg simvastatin. AICAR, 5-aminoimidazole-4-carboxamide ribonucleoside; DMSO, dimethyl sulfoxide;VO2, oxygen consumption; WT, wild type; YS, Y524S.
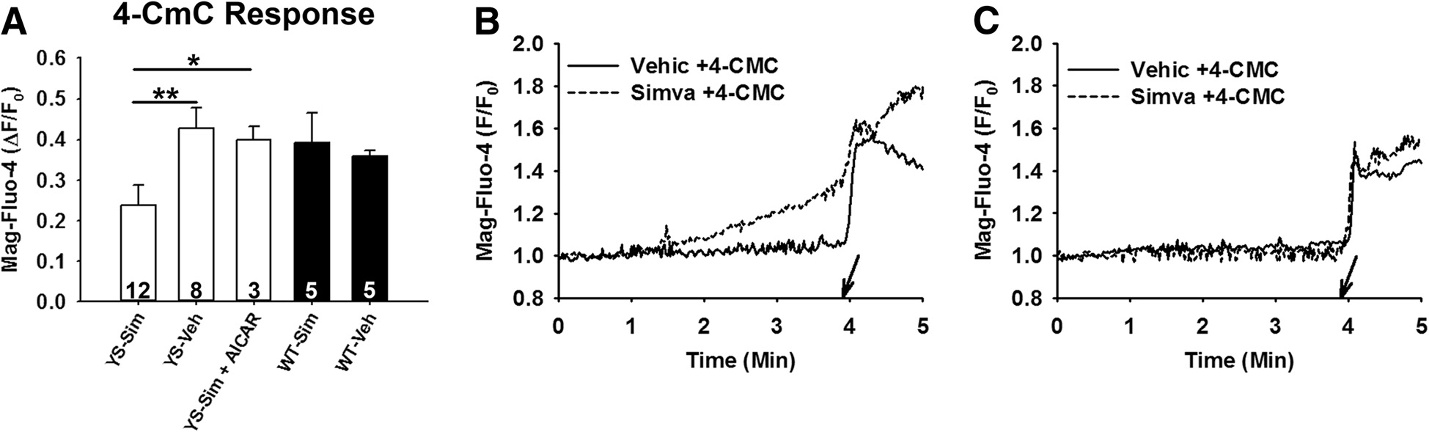
Simvastatin depletes SR Ca2+ stores in FDB fibers isolatedfrom YS miceCa2+ stores in YS FDB fibers are decreased by exposureto elevated temperatures To determine if a reduc-tion in Ca2+ stores occurs with simvastatin, we usedMag-fluo-4, a low-affinity Ca2+ indicator, and 4-CmC toassess the readily releasable SR Ca2+ stores [4-CmCwas applied to isolated fibers immediately after a 3-minute incubation with simvastatin. We found a signifi-cant (P <0.05) decrease in the readily releasable Ca2+stores in YS fibers exposed to 500 μM simvastatin com-pared with YS fibers exposed to vehicle-only (Figure while no difference was found in WT fibers at this con-centration of simvastatin. This finding suggests that theincreased cytosolic Ca2+ levels in the YS mice that occurafter exposure to simvastatin are due to SR Ca2+ leakleading to SR Ca2+ store depletion.
Figure 2 Isolated fibers from YS mice exhibit increased
We assessed the ability of AICAR to regulate the
sensitivity to simvastatin compared to WT mice. Dose–response
simvastatin-induced increase in Ca2+ leak in the YS fi-
curves from isolated WT and YS FDB fibers incubated for 3 minutes
bers. Isolated YS fibers were incubated with 1 mM
in respective doses of simvastatin. Data points reflect peak cytosolic
AICAR prior to incubation with 500 μM simvastatin. As
Ca2+ change from baseline, indicating that fibers from YS micerespond to simvastatin at lower doses than WT fibers. Fibers were
shown in Figure , Ca2+ stores were protected from
used only at a single simvastatin concentration. Each data point
the simvastatin-induced depletion by prior administra-
represents the mean cytosolic Ca2+ response from a minimum of
tion of AICAR (P <0.01). We determined if AICAR
three fibers taken from three separate mice. Ca2+, calcium; FDB,
could also prevent the simvastatin-induced Ca2+ release
flexor digitorum brevis; WT, wild type; YS, Y524S.
at higher simvastatin doses in WT mice (Figure
Knoblauch et al. Skeletal Muscle 2013, 3:22
Figure 3 Pre-incubation with simvastatin decreases the 4-CmC-modulated cytosolic Ca2+response in isolated FDB fibers from YScompared to WT mice. Represented as the change (Δ) from baseline to peak values, (A) shows that upon exposure to 1 mM 4-CmC thoseYS fibers incubated for 3 minutes in 500 μM simvastatin (YS-Sim) release significantly less Ca2+ from the SR than YS fibers receiving vehicle-only(YS-Veh) incubation (P <0.01) and from YS fibers incubated in 1 mM AICAR followed by 500 μM simvastatin (YS-Sim + AICAR) (P <0.05). Numbersrepresent total fibers used per group from a minimum of three mice. (B) YS and (C) WT show representative Mag-fluo-4 fluorescencetracings in single fibers exposed to either simvastatin or vehicle. Arrows indicate the time point at which 4-CmC was applied to thefibers. AICAR, 5-aminoimidazole-4-carboxamide ribonucleoside; 4-CmC, 4-chloro-m-cresol; Ca2+, calcium; FDB, flexor digitorum brevis; SR,sarcoplasmic reticulum; WT, wild type; YS, Y524S.
When WT fibers were incubated with 1 mM simvastatin,
we found that AICAR pre-treatment also greatly de-
Despite the prevalence of statin myopathies, a mechan-
creased Ca2+ release in WT fibers (P <0.001), suggesting
ism to explain the underlying trigger has remained elu-
that statins have the potential to trigger Ca2+ release in
sive. The current study's objective was to determine
normal fibers but require higher simvastatin concentra-
whether a MH-associated defect in RyR1 increased sen-
tions than YS fibers and that AICAR may be a useful
sitivity to simvastatin and whether AICAR, which pre-
intervention for SIM even in patients without RyR1
vents heat-induced HMR in the YS mice, blocked the
response to simvastatin. We show that the YS mice
Figure 4 Pre-treatment with AICAR reduces the cytosolic Ca2+ response to simvastatin. Pre-incubation with AICAR prevents Ca2+ releasein (A) YS fibers exposed to 500 μM simvastatin (P <0.01) and in (B) WT fibers exposed to 1 mM simvastatin (P <0.001). (C) YS and (D) WT showchange in Ca2+ concentration for AICAR-treated (dashed) and untreated (solid) fibers after exposure to either 500 μM (YS) or 1 mM (WT)simvastatin at 2 minutes . AICAR, 5-aminoimidazole-4-carboxamide ribonucleoside; Ca2+, calcium; WT, wild type; YS, Y524S.
Knoblauch et al. Skeletal Muscle 2013, 3:22
display an MH-like response (elevated VO2, sustained
lower simvastatin concentrations in the presence of a
muscle contractures, elevated body temperature) to an
RyR1 mutation associated with MH in humans. The
acute dose of simvastatin, and the degree of response is
prevalence of genetic abnormalities capable of causing
dose-dependent. Simvastatin also enhances SR Ca2+ leak
MH has been estimated to be as low as 1:3,000
and SR Ca2+ store depletion in FDB fibers from both YS
Whereas the incidence of SIM is relatively low (approxi-
and WT mice but the response in WT mice requires
mately 10%) among the millions of statin users, it is
higher concentrations of simvastatin. In FDB fibers from
highly possible that those individuals exhibiting signs
both YS and WT mice, the response to simvastatin was
and symptoms of SIM are harboring an underlying RyR1
prevented by AICAR, suggesting that even in WT fibers
myopathy. Guis et al. showed that seven of nine individ-
the effect of simvastatin involves RyR1.
uals exhibiting symptoms of severe statin myopathy were
AICAR is a known activator of the energy sensing kin-
found to have a positive IVCT, indicative of an under-
ase, AMP-activated protein kinase (AMPK). We recently
lying RyR1 abnormality . Therefore, further research
demonstrated, however, that AICAR also has a direct ef-
is needed to determine whether individuals experiencing
fect on RyR1 and rescues the YS mice from heat-induced
SIM also have mutations in RyR1. If true, drugs such as
sudden death independent of AMPK activation []. We
AICAR that modulate RyR1 activity can be investigated
now demonstrate that treatment of YS mice with AICAR,
as a potential therapy for these individuals, which may
which decreases Ca2+ leak in the presence of cellular levels
allow continued statin use without the side effects asso-
of ATP [], prevents the simvastatin-associated increases
ciated with SIM.
in VO2 and heat production as well as greatly attenuatesCa2+ leak from the SR upon exposure of FDB fibers to
simvastatin. AICAR also largely eliminates the statin-
The YS mutation in RyR1 increases the sensitivity to the
induced Ca2+ release in healthy WT mice. These results
cholesterol-lowering medication simvastatin. This sensi-
suggest that AICAR might also be a potential therapeutic
tivity is marked by systemic increases in VO2, muscle
intervention to prevent statin myopathies associated with
contractures and heat production due to a temporal re-
RyR1 mutations in sensitive individuals and protect
lease of Ca2+ into the cytosol from the SR. Pharmaceutical
against myopathies arising from high statin doses in indi-
interventions that decrease Ca2+ leak from RyR1 (such as
viduals without RyR1 mutations.
AICAR) prevent both the systemic manifestation of SIMand the statin-induced Ca2+ release from the SR in single
YS mutation explains clinical symptoms of SIM
fibers. We show that RyR1 mutation increases sensitivity
Alterations in Ca2+ signaling with simvastatin could ex-
to SIM, suggesting that individuals affected by SIM could
plain many of the symptoms associated with SIM in
harbor underlying RyR1 mutations and that AICAR may
humans including muscle fatigue, cramping, and increased
be an effective therapeutic intervention.
levels of circulating CK. Depletion of stores contributes tofatigue, while increased resting Ca2+ is known to trigger
Ca2+ release and muscle contraction, giving rise to muscle
4-CmC: 4-chloro-m-cresol; AICAR: 5-aminoimidazole-4-carboxamide
cramping similar to that which occurs in Brody disease.
ribonucleoside; AMPK: AMP-activated protein kinase; BTS: 4-methyl-N-(phenylmethyl)benzenesulfonamide; Ca2+: Calcium; CK: Creatine kinase;
Brody disease results from a reduction in the number
DMEM: Dulbecco's modified Eagle's medium; DMSO: Dimethyl sulfoxide;
and activity of sarco/endoplasmic reticulum Ca2+-ATPase
EC50: Half maximal effective concentration; EDTA: Ethylenediaminetetraacetic
(SERCA) proteins in skeletal muscle, which inhibits the
acid; EtOH: Ethanol; FDB: flexor digitorum brevis; Fura-2 AM: Fura-2acetoxymethyl ester; HMG-CoA: 3-hydroxy-3-methylglutaryl coenzyme-A;
re-uptake of cytosolic Ca2+ during muscle activity ,].
HMR: Hypermetabolic response; IP: Intraperitoneal; IVCT: In vitro contracture
Individuals afflicted with Brody disease complain of fa-
test; LDL: Low-density lipoprotein; MH: Malignant hyperthermia;
tigue as well as muscle cramping that is exacerbated dur-
RyR1: Ryanodine receptor type 1; SERCA: Sarco/endoplasmic reticulum Ca2+−ATPase; SIM: Statin-induced myopathy; SR: Sarcoplasmic reticulum;
ing periods of increased activity such as exercise [].
VO2: Oxygen consumption; VO2 max: Maximum oxygen consumption;
These symptoms reflect those commonly reported among
WT: Wild type; YS: Y524S.
individuals experiencing SIM. Separately, elevated circu-lating CK levels among individuals experiencing SIM can
Competing interests
also be explained by rhabdomyolysis triggered by the
The authors declare that they have no competing interests.
statin-modulated increase in cytosolic Ca2+ levels and acti-vation of calpains Elevated CK levels are com-
Authors' contributionsMK conceived and developed the study, conducted indirect calorimetry
monly experienced by individuals experiencing SIM.
experiments, assisted with Ca2+ imaging experiments, performed statisticalanalyses, and prepared the draft manuscript. ADA conducted Ca2+ imaging
Clinical relevance
experiments, assisted with analyses, prepared data, and assisted withmanuscript preparation. SLH created and maintained the YS mouse line,
An acute dose of simvastatin increases cytosolic Ca2+
assisted with study design and data interpretation, and assisted with
levels within the myofiber and this increase occurs at
manuscript preparation. All authors read and approved the final manuscript.
Knoblauch et al. Skeletal Muscle 2013, 3:22
G, van Engelen B: Brody syndrome: a clinically heterogeneous entity
This was supported by NIH grants 5R01AR041802 and 5R01AR053349 to SLH.
distinct from Brody disease: a review of literature and a cross-sectional
A.D.A. was supported by a postdoctoral fellowship from the Mexican Council
clinical study in 17 patients. Neuromuscul Disord 2012, 22:944–954.
of Science and Technology (186607). We would like to thank the Mouse
Belcastro A, Shewchuk L, Raj D: Exercise-induced muscle injury: a calpain
Phenotyping Core at Baylor College of Medicine, Houston, TX, USA, for the
hypothesis. Mol Cell Biochem 1998, 179(1–2):135–145.
use and assistance with the Oxymax indirect calorimetry system.
Raastad T, Owe S, Paulsen G, Enns D, Overgaard K, Crameri R, Kiil S,Belcastro A, Bergersen L, Hallén J: Changes in calpain activity, muscle
Received: 9 April 2013 Accepted: 9 August 2013
structure, and function after eccentric exercise. Med Sci Sports Exerc 2010,
Published: 3 September 2013
Cite this article as: Knoblauch et al.: Mice with RyR1 mutation (Y524S)
Joy T, Hegele R: Narrative review: statin-related myopathy. Ann Intern Med
undergo hypermetabolic response to simvastatin. Skeletal Muscle
Sathasivam S, Lecky B: Statin induced myopathy. BMJ 2008, 337:1159–1162.
Venero C, Thompson P: Managing statin myopathy. Endocrinol Metab ClinNorth Am 2009, 38(1):121–136.
Ghatak A, Faheem O, Thompson PD: The genetics of statin-inducedmyopathy. Atherosclerosis 2010, 210(2):337–343.
Sirvent P, Fabre O, Bordenave S, Hillaire-Buys D, Raynaud De Mauverger E,Lacampagne A, Mercier J: Muscle mitochondrial metabolism and calciumsignaling impairment in patients treated with statins. Toxicol ApplPharmacol 2012, 259(2):263–268.
Sirvent P, Mercier J, Lacampagne A: New insights into mechanisms ofstatin-associated myotoxicity. Curr Opin Pharmacol 2008, 8(3):333–338.
Sirvent P, Bordenave S, Vermaelen M, Roels B, Vassort G, Mercier J, Raynaud E,Lacampagne A: Simvastatin induces impairment in skeletal muscle whileheart is protected. Biochem Biophys Res Commun 2005, 338(3):1426–1434.
Sirvent P, Mercier J, Vassort G, Lacampagne A: Simvastatin triggersmitochondria-induced Ca2+ signaling alteration in skeletal muscle.
Biochem Biophys Res Commun 2005, 329(3):1067–1075.
Inoue R, Tanabe M, Kono K, Maruyama K, Ikemoto T, Endo M: Ca2+-releasingeffect of cerivastatin on the sarcoplasmic reticulum of mouse and ratskeletal muscle fibers. J Pharmacol Sci 2003, 93(3):279–288.
Metterlein T, Schuster F, Tadda L, Hager M, Roewer N, Anetseder M: Statinsalter intracellular calcium homeostasis in malignant hyperthermiasusceptible individuals. Cardiovasc Ther 2010, 28(6):356–360.
Guis S, Figarella-Branger D, Mattei JP, Nicoli F, Le Fur Y, Kozak-Ribbens G,Pellissier JF, Cozzone PJ, Amabile N, Bendahan D: In vivo and in vitrocharacterization of skeletal muscle metabolism in patients with statin-induced adverse effects. Arthritis Rheum 2006, 55(4):551–557.
Rosenberg H, Davis M, James D, Pollock N, Stowell K: Malignanthyperthermia. Orphanet J Rare Dis 2007, 2:21.
Chelu M, Goonasekera S, Durham W, Tang W, Lueck J, Riehl J, Pessah I,Zhang P, Bhattacharjee M, Dirksen R, Hamilton SL: Heat- and anesthesia-induced malignant hyperthermia in an RyR1 knock-in mouse. FASEB J2006, 20:329–330.
Leung B, Sattar N, Crilly A, Prach M, McCarey D, Payne H, Madhok R,Campbell C, Gracie J, Liew F, McInnes I: A novel anti-inflammatory role forsimvastatin in inflammatory arthritis. J Immunol 2003, 170:1524–1530.
Lanner JT, Georgiou DK, Dagnino-Acosta A, Ainbinder A, Cheng Q, Joshi AD,Chen Z, Yarotskyy V, Oakes JM, Lee CS, Monroe TO, Santillan A, Dong K,Goodyear L, Ismailov II, Rodney GG, Dirksen RT, Hamilton SL: AICARprevents heat-induced sudden death in RyR1 mutant mice independentof AMPK activation. Nat Med 2012, 18(2):244–251.
Dagnino-Acosta A, Guerrero-Hernández A: Variable luminal sarcoplasmicreticulum Ca2+buffer capacity in smooth muscle cells. Cell Calcium 2009,46:188–196.
Durham W, Aracena-Parks P, Long C, Rossi A, Goonasekera S, Boncompagni
Submit your next manuscript to BioMed Central
S, Galvan D, Gilman C, Baker M, Shirokova N, Protasi F, Dirksen R, Hamilton
and take full advantage of:
S: RyR1 S-nitrosylation underlies environmental heat stroke and suddendeath in Y522S RyR1 knockin mice. Cell 2008, 133:53–65.
• Convenient online submission
Herrmann-Frank A, Richter M, Lehmann-Horn F: 4-Chloro-m-cresol: aspecific tool to distinguish between malignant hyperthermia-susceptible
• Thorough peer review
and normal muscle. Biochem Pharmacol 1996, 52:149–155.
• No space constraints or color figure charges
Vattemi G, Gualandi F, Oosterhof A, Marini M, Tonin P, Rimessi P, Neri M,
• Immediate publication on acceptance
Guglielmi V, Russignan A, Poli C, van Kuppevelt T, Ferlini A, Tomelleri G:Brody disease: insights into biochemical features of SERCA1 and
• Inclusion in PubMed, CAS, Scopus and Google Scholar
identification of a novel mutation. J Neuropathol Exp Neurol 2010,
• Research which is freely available for redistribution
Voermans N, Laan A, Oosterhof A, van Kuppevelt T, Drost G, Lammens M,Kamsteeg E, Scotton C, Gualandi F, Guglielmi V, van den Heuvel L, Vattemi
Submit your manuscript at www.biomedcentral.com/submit
Source: http://www.skeletalmusclejournal.com/content/pdf/2044-5040-3-22.pdf
19_kotler_app.qxd 1/18/07 4:51 PM Page A18 Appendix 2 CBC Video Cases Each video case corresponds to specific chapters. See the correlation table below fordetails. Video Case 1: Chapters 1, 2, 3Video Case 2: Chapters 4, 6, 8Video Case 3: Chapters 4, 5, 16Video Case 4: Chapters 8, 9Video Case 5: Chapters 4, 11, 13Video Case 6: Chapters 10, 16Video Case 7: Chapter 14, 15
Estás de corazón Red Hispanoportuguesa de Espiritualidad Apostólica Marista A la memoria de Servando, Julio, Miguel Angel y Fernando que con su gesto de entrega total a los más pobres, hasta llegar al martirio, nos enseñaron cómo se unifica la vida en el amor, cómo se adora y sirve a Dios en la vida cotidiana y cómo ésta se hace presente